
Dendroclimatic analysis of white pine(Pinusstrobus L.)using long-term provenance test sites across eastern North America
2018-09-12 09:14:42SophanChhinRonaldZalesnyJrWilliamParkerandJohnBrissette
Forest Ecosystems 2018年3期
Sophan Chhin,Ronald S.Zalesny Jr,William C.Parker and John Brissette
Abstract Background:The main objective of this study wasto examine the climatic sensitivity of the radial growth response of 13 eastern white pine(Pinusstrobus L.)provenances planted at seven test sites throughout the northern part of the species’native distribution in eastern North America.Methods:The test sites(i.e.,Wabeno,Wisconsin,USA;Manistique,Michigan,USA;Pine River,Michigan,USA;Newaygo,Michigan,USA;Turkey Point,Ontario,Canada;Ganaraska,Ontario,Canada;and Orono,Maine,USA)examined in this study were part of arange-wide white pine provenance trial established in the early 1960sin the eastern United States and Canada.Principal componentsanalysis(PCA)wasused to examine the main modesof variation[first(PC1)and second(PC2)principal component axes]in the standardized radial growth indicesof the provenancesat each test site.The year scoresfor PC1 and PC2 were examined in relation to an array of test site climate variablesusing multiple regression analysisto examine the commonality of growth response acrossall provenancesto the climate of each test site.Provenance loadingson PC1 and PC2 were correlated with geographic parameters(i.e.,latitude,longitude,elevation)and a suite of biophysical parametersassociated with provenance origin location.Results:The amount of variation in radialgrowth explained by PC1 and PC2 ranged from 43.4%to 89.6%.Dendroclimatic modelsrevealed that white pine radial growth responsesto climate were complex and differed among sites.The key dendroclimatic relationshipsobserved included sensitivity to high temperature in winter and summer,cold temperature in the spring and fall(i.e.,beginning and end of the growing season),summer moisture stress,potentialsensitivity to storminduced damage in spring and fall,and both positive and negative effectsof higher winter snowfall.Separation of the loadingsof provenanceson principal component axeswasmainly associated with temperature-related bioclimatic parametersof provenance origin at 5 of the 7 test sitesclose to the climate influence of the Great Lakes(i.e.,Wabeno,Manistique,Pine River,Newaygo,and Turkey Point).In contrast,differencesin radialgrowth response to climate at the Ganaraskatest site,were driven more by precipitation-related bioclimatic parametersof the provenance origin location while radial growth at the easternmost Orono test site wasindependent of bioclimate at the provenance origin location.Conclusions:Study resultssuggest that genetic adaptation to temperature and precipitation regime may significantly influence radial growth performance of white pine populationsselected for use in assisted migration programsto better adapt white pine to a future climate.
Keywords:Adaptation,Climate change,Dendrochronology,Seed source
Background
If fossil fuel intensive energy generation continues to grow at current rates,atmospheric concentrations of carbon dioxide(CO2)and other greenhouse gases will increase dramatically.Continued high rates of emissions over the next century could raise mean annual global surfacetemperatureby 2.5°C to 6.5°C by 2100(IPCC 2007).The resulting projected change in spatial and temporal temperatureand precipitation regimes,increased frequency of natural disturbance events,and atmospheric CO2enrichment isexpected to alter thedistribution,structure,function,and productivity of northern forests(Mickler et al.2000;Parmesan and Yohe 2003).
A critical need exists to develop ecologically sustainable forest management approaches that incorporate climate change adaptation to sustain carbon sequestration,fiber production,and other ecological goodsand servicesprovided by managed forests(Millar et al.2007;Alexander and Perschel 2009).The success of such efforts depends in part on knowledge of genetic variation in climatic response of tree species(Rehfeldt et al.1999;Millar et al.2007;Wang et al.2010).Therefore,information generated from long-term provenance testshassubstantial potential to improveunderstanding of tree species responses and adaptability to a rapidly changing climate(Rehfeldt et al.1999;Wang et al.2010).
Eastern white pine(Pinus strobus L.)is an ecologically and economically important softwood species that occurs throughout much of eastern North America(Wendel and Smith 1990).White pine was a dominant component of pre-settlement forests of eastern North America that is highly valued for its exceptional wood properties.The effects of large-scale exploitive logging and catastrophic fire in the 18th and 19th centuries,coupled with more recent institutional fire suppression and damage and mortality from white pine blister rust(Cronartium ribicola J.C.Fisch.),white pine weevil(Pissodes strobi(Peck)),and root rot fungi(Armillaria mellea(Vahl)P.Kumm)have dramatically reduced the abundance of white pine on the landscape(Abrams et al.2000).
Genetic adaptation refers to genetic differences or variation among provenances in the natural range of a tree species that have evolved over several generations and ensure it is in sync with the environmental conditions of its location(White et al.2007;King et al.2013;Gray et al.2016).Examples of genetic adaptation in trees include the link between provenance location and tree growth rate and phenology,i.e.,tree provenances from more northern latitudes generally grow more slowly and start growing earlier in spring/stop earlier in fall compared to those from more southern latitudes.White pine typically exhibits significant adaptive genetic variation in response to broad climatic gradients,the product of natural selection and genetic processes(Wright 1970;Genys 1987),and as a result is found as a component of most upland forest types in its range.This variation has resulted in tree species populations that are genetically adapted to local and regional environmental conditions.However,as the climate continues to change,white pine may become increasingly unsynchronized with and maladapted to the prevailing climate,resulting in decreased vigor,productivity,and wood quality.The geographic shifts and reductions in suitable climatic habitat projected by climate change models(McKenney et al.2007;Iverson et al.2008;Joyce and Rehfeldt 2013)represent a serious threat to white pine throughout much of eastern North America,especially as changes in climate will also influence other abiotic and biotic stressors(Millar et al.2007).
A network of provenance test sites established in the early 1960s has provided insight into the adaptive genetic variation of white pine(Wright 1970;Genys 1987;Joyce and Rehfeldt 2013;Zalesny and Headlee 2015).This scientific resource has renewed value as a long-term experiment that can increase understanding of the effect of climate change on white pine tree populations growing outside the climate envelope to which they are naturally adapted(Wang et al.2006).For example,it is known that southern sources of white pine grow faster but are more susceptible to frost damage when planted farther north(Joyce and Rehfeldt 2013).Combining such knowledge with constantly improving global climate models and climate change scenarios can inform seed deployment decisions to produce nursery stock that is best adapted to a given planting site(Joyce and Rehfeldt 2013;Yang et al.2015).In addition,such knowledge can guide management of natural regeneration where local provenancesare currently well adapted to projected future climate conditions.
Dendrochronological methods can be effectively applied to describe the genetic adaptation of tree provenances to environmental conditions(e.g.,Chhin 2010;McClane et al.2011;Pluess and Weber 2012;King et al.2013;Eilmann et al.2014;Chhin 2015)and to examine geographic variation in response to climate(e.g.,Chen et al.2010).In response to seasonal changes in climatic drivers of tree phenology,the annual growth cycle of temperate and boreal tree speciesin North Americaalternatesbetween phases of summer growth and winter dormancy(Fritts 1976).Examination of the seasonal periodicity of climatic effects on radial stem growth processes through tree-ring analysis(dendrochronology)provides insight into the seasonal timing of radial growth–climate relationships(Fritts 1976;Chhin et al.2008)and therefore can be used to quantify genetic adaption of tree provenancesto environmental conditions(Chhin 2010;Pluessand Weber 2012;Chhin 2015).
The primary goal of this study was to identify eastern white pine provenanceswith enhanced adaptation to climate changethroughout theportion of thespecies’nativedistribution from Wisconsin to Maine,USA,and including Ontario and Quebec,Canada.Selection of climatically adapted provenances for planting will help promote biologically and economically sustainable reforestation,afforestation,and gene conservation throughout the region.While genetic tree improvement studies have focused on optimizing selection of growth traits such as maximizing tree height(Joyce et al.2002;Lu et al.2003b),few studies have examined the effect of climatic sensitivity on these growth traits(Lu et al.2003a;Chhin 2008;Joyce and Rehfeldt 2013;Chhin 2015).Using growth-climate models and data collected from long-term white pine provenance tests,the specific objectives of this study wereto use adendroclimatological approach to:
1)Examinethedegreeof similarity in climatic sensitivity among theprovenancesplanted at each of the7 test sites.
2)Examine the influence of geographic location and bioclimate of provenance origin on radial growth response at each test sites.
Methods
The study was conducted at 7 field test sites belonging to a range-wide white pine provenance trial established in the early 1960s in the eastern United States and Canada(Wright et al.1970)(Fig.1;Table 1).Thirteen white pine provenances were evaluated at Wabeno,Wisconsin,USA;Manistique,Michigan,USA;and Pine River,Michigan,USA(Table 2).The Penobscot,Maine,USA provenance was absent at the Newaygo,Michigan,USA test site.The Newaygo provenance was not tested at Turkey Point,Ontario,Canada;Ganaraska,Ontario,Canada;and Orono,Maine,USA test sites.The field sampling design varied at each test site,with a uniform spacing of 2.13 m×2.13 m.Thefield design at the Wabeno,Manistique,and Pine River test sites were planted in 24 blocks with one tree per provenance per block.At the Newaygo and Ganaraska test sites,white pine was planted in 4 blocks comprising 81 trees per provenance(arranged in 9 rows of 9 trees)per block.The Turkey Point test site consisted of 4 blocks,each block containing 11 rows of 11 trees per provenance.At the Orono test site,white pine was planted in 12 blocks and each block contained each provenance planted in 4-tree row plots(King and Nienstaedt 1969;Zalesny and Headlee2015).
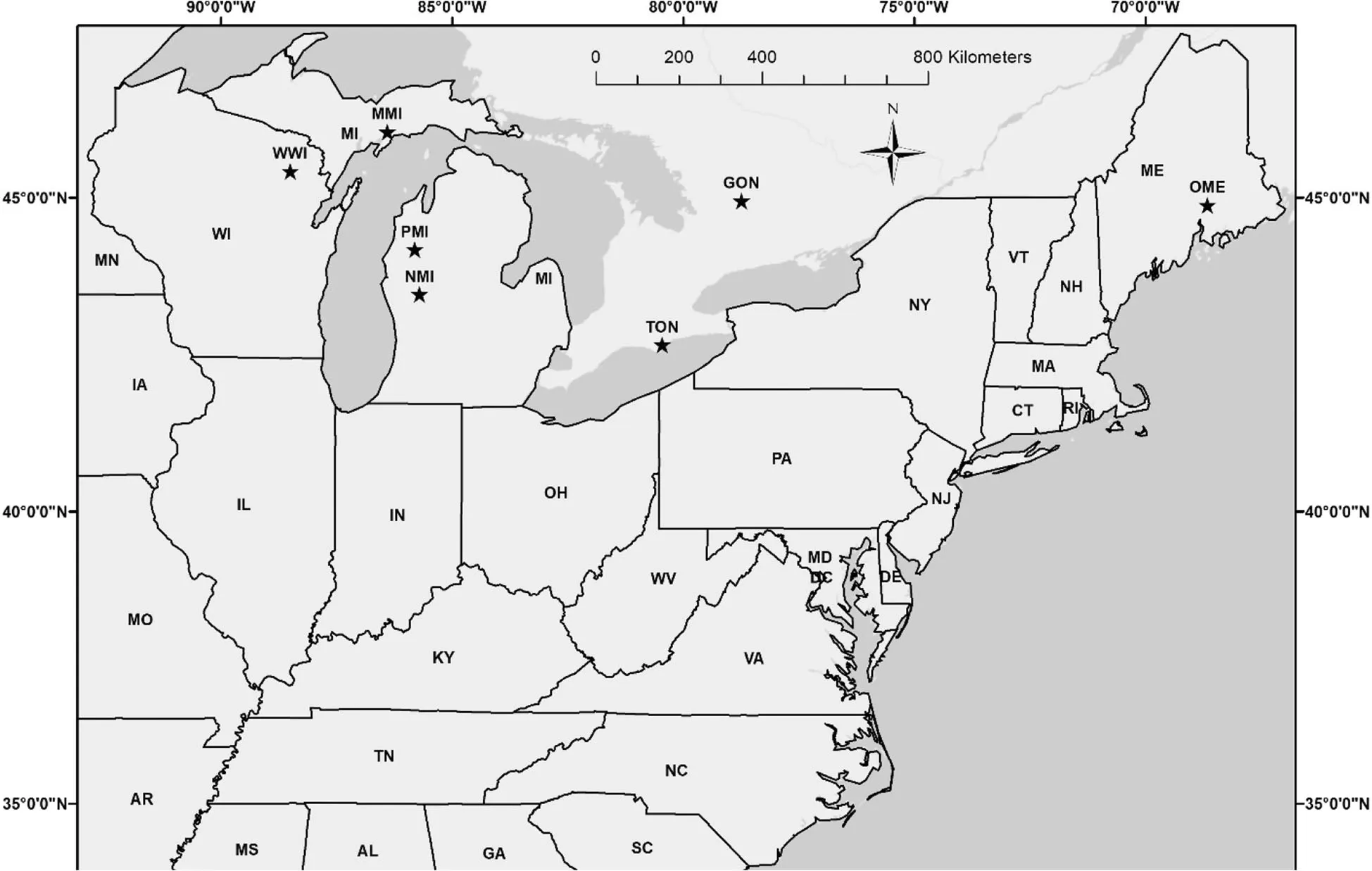
Fig.1 Distribution of the white pine provenance trial test sites(denoted by stars)in the United Statesand Canadathat were assessed in thisstudy.WWI=Wabeno,WI;MMI=Manistique,MI;PMI=Pine River,MI;NMI=Newaygo,MI;TON=Turkey Point,ON;GON=Ganaraska,ON;OME=Orono,ME
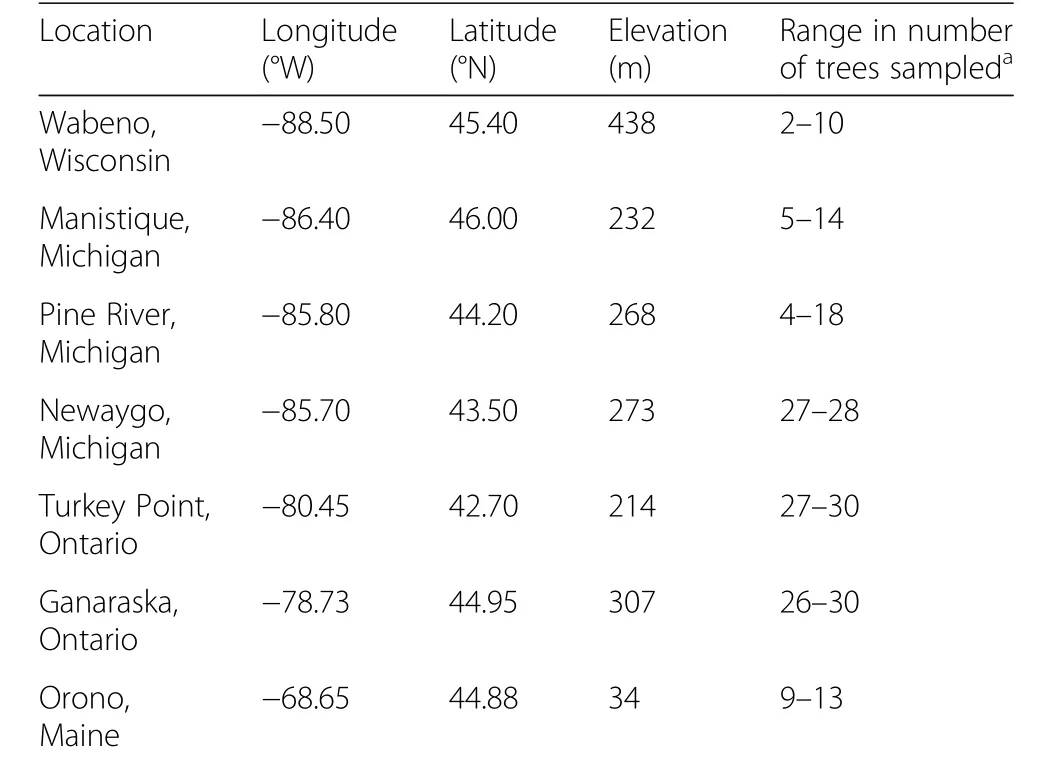
Table 1 Location,longitude,latitude,elevation(meters(m))and number of trees sampled for the 7 test sites
Two increment cores were collected at breast height(i.e.,1.37 m)from a subset of trees that had a diameter at breast height(dbh)at or near the median value of all living trees measured in a plot.All increment cores used in this study were taken at dbh in the east cardinal direction using a 5-mm diameter borer.The number of living trees per provenance available for sampling(Table 1)differed due to variable tree mortality among test sites.The cores were permanently mounted and sanded using standard dendrochronology procedures to prepare them for radial growth trend analysis(Stokes and Smiley 1996).Scanned images of individual cores were acquired with a flatbed scanner at a resolution of 720 dpi.Cores and scanned images were visually crossdated.Ring-widths were measured from the scanned images using an image analysis system(WinDENDRO,Regent Instruments,Québec,Canada).Statistical quality control was conducted using COFECHA software to ensure that missing and false rings were accounted for(Holmes 1983;Grissino-Mayer 2001).To remove non-climatic effects,cross-dated ring-width measurementswerede-trended using aconservative de-trending approach that included fitting a negative exponential curve,simple linear regression,and a line through the mean with ARSTAN software(Cook 1985).The standardized version of the chronology was retained for each individual core sample to prevent the loss of any radial growth signals associated with provenances at different test sites.The standardized radial growth indices were then averaged for individual trees of each provenance at each test site.Chronology statistics,including mean sensitivity and the inter-series correlation coefficient,were determined for each provenancelocation at each test site.
Principal components analysis(PCA)wasused to examine the main modes of variation in the standardized radial growth indices of individual provenances at each test site(Graumlich 1993;Legendre and Legendre 1998).The PCA was carried out using the Factor Analysis function in Systat(ver.13.0,released 2009),which also included selection of a Varimax rotation to improve interpretation of the principal component axes.Selected principal component axes explained more variation than expected,based on a broken stick null model(Legendre and Legendre 1998).
Year scores from 1976 to 2009 were used in dendroclimatic analyses to examine shared provenance radial growth responses to test site climate.For test sites in the United States,primary climate data were obtained from PRISM(Daly 2008)and for test sites in Canada were obtained from ClimateNA(Hamann et al.2013).Climate data included minimum,maximum,and mean monthly temperature and total monthly precipitation.These primary variables were used as the basis to produce a synthetic monthly climatic moisture index(CMI)variable calculated as total monthly precipitation minus a measure of monthly potential evapotranspiration(Hogg 1997).Since tree radial growth may respond more strongly when climatic variables are expressed at a seasonal compared to a monthly scale,monthly climatic variables were converted to seasonal 3-month periods(i.e.,quarters)by averaging across months for temperature-related variables and summing across months for precipitation and moisture index variables.Because climatic conditions in the previous year can influence radial growth in the following growing season,the range of monthly and seasonal periods was examined from April of the prior year(t-1)to October of the current year(t)of ring formation.
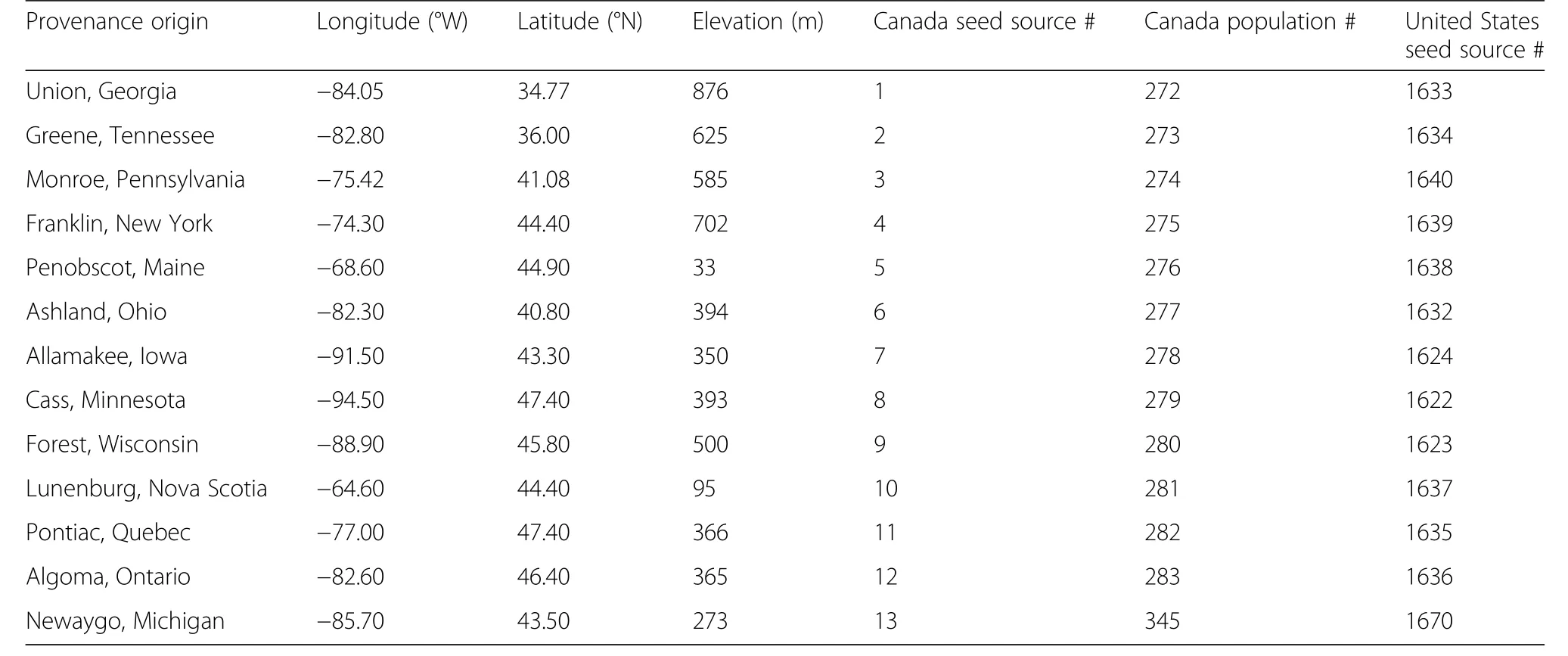
Table 2 Location,longitude,latitude,elevation(meters(m)),and seed source identification numbers for the 13 provenances tested at 7 test sites
Dendroclimatic analysis was conducted to examine the relationship between year scores representing the main modes of variation(i.e.,the first(PC1)and second(PC2)principal component axes)shared across provenances at each test site with respect to the monthly and seasonal climate at each test site.Dendroclimatic analysis was conducted using a step-wise multiple regression conducted with R statistical analysis software for the period 1976 to 2009(Venablesand Ripley 2002;Chhin et al.2008;RCore Team 2014).Standardized partial regression coefficients were also calculated to rank the relative importance of the predictor variables in each regression model(Zar 1999).
The loadings of each provenance on the significant principal component axes were used to examine the influence of the bioclimate of the provenance origin location on radial growth responsesat the7 test sites.A suite of 19 bioclimatic parameters for the climate normal period of 1971 to 2000 were obtained from McKenney et al.(2007)(Table 3).The loadings of each provenance on PC1 and PC2 of each test site were correlated with geographical parameters(i.e.,latitude,longitude,elevation,and distance from test site)of each provenance origin location.Sensitivity of the loadings of each provenance on PC1 and PC2 were therefore also examined in relation to the 19 bioclimatic parameters(Table 3).Given the low number of provenances at a site(n=12 or 13 provenances),all correlation analyses were conducted using the Spearman rank method using Systat(ver.13.0,released 2009).
Results
Chronology statistics,including mean sensitivity and the inter-series correlation coefficients,are provided in Additional file 1.Mean sensitivity of provenances was significantly correlated with latitude only at the Newaygo test site(ρ=0.53)(results not presented).The variance in radial growth among white pine provenances captured by PC1 was statistically significant for all test sites(Table 4).The amount of explained variance was highest for Ganaraska(58.8%)and lowest for Wabeno(31.9%).All of the second principal component axes were significant,except for Wabeno.
Shared provenance radial growth responses to climate of each test site
Significant linear relationships were exhibited between PCI and PC2 scores and the monthly and seasonal temperature and precipitation regimesat several test sites.The year scores of whitepineradial growth with respect to PC1(Fig.2a)were negatively associated with mean maximum temperature forthe winter seasonal period at Manistique and Newaygo[December(t-1)to February(t)],and Orono[January(t)to March(t)](Fig.3).The year scores on PC1 at Pine River were negatively related to the summer seasonal maximum temperature[May(t)to July(t)].At Orono,radial growth also responded positively to the fall seasonal period of maximum temperature in the previous growing season[September(t-1)to November(t-1)].White pine radial growth at Wabeno,Turkey Point,and Ganaraskawasnot responsiveto mean maximum temperature.Relationshipsbetween theyear scores of PC2(Fig.2b)and maximum temperature indicated that whitepineradial growth wasnegatively related with September(t)monthly maximum temperature at Newaygo and Turkey Point(Fig.3),and was negatively related to summer(t)temperature at Ganaraska and Orono.The PC2 year scores for Wabeno and Orono indicated that growth was positively associated with spring maximum temperature.The PC2 year scores were also positively related with September(t-1)maximum temperaturein Manistique.
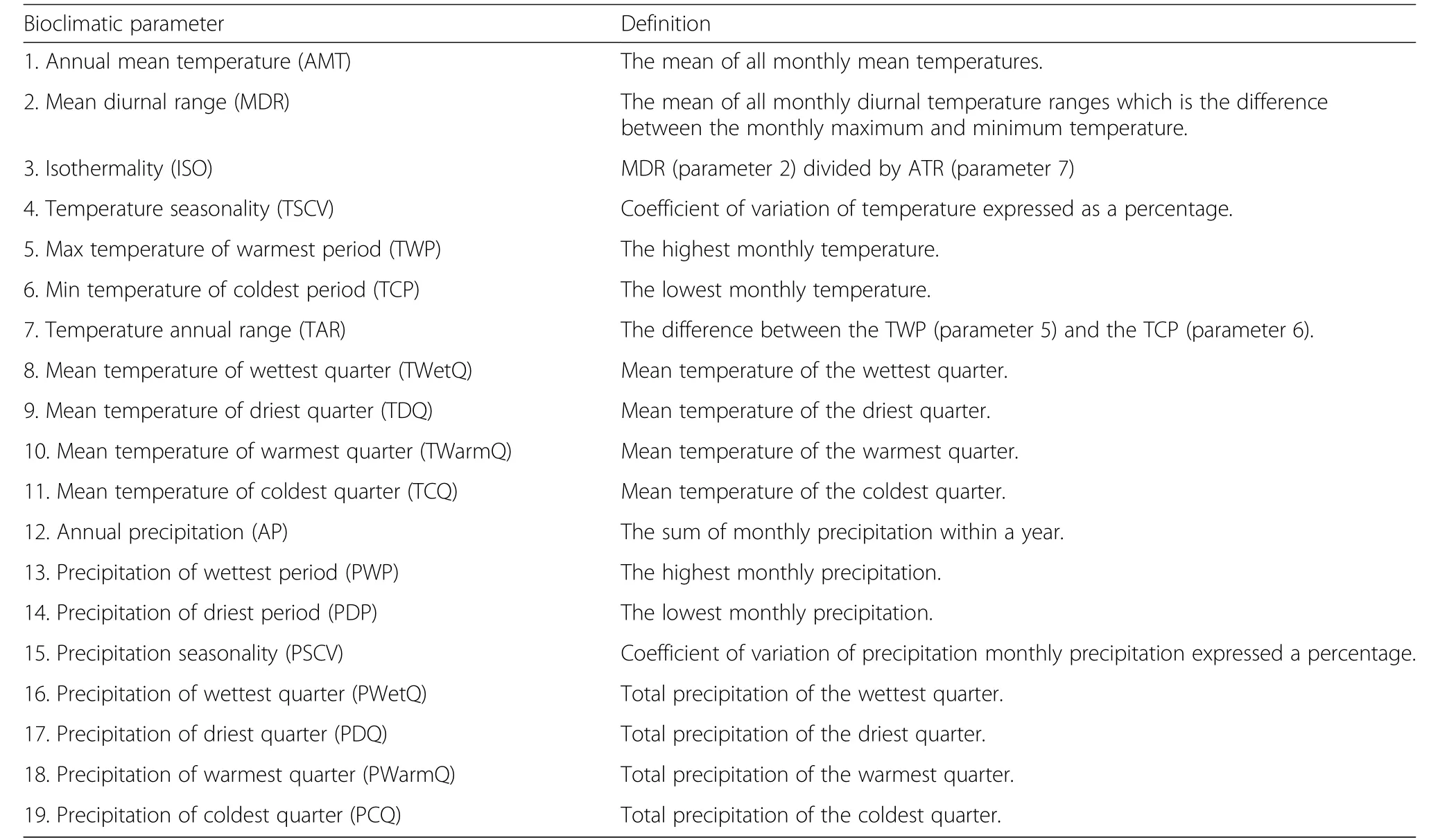
Table 3 Bioclimatic parameters(abbreviations)and their definitionsa
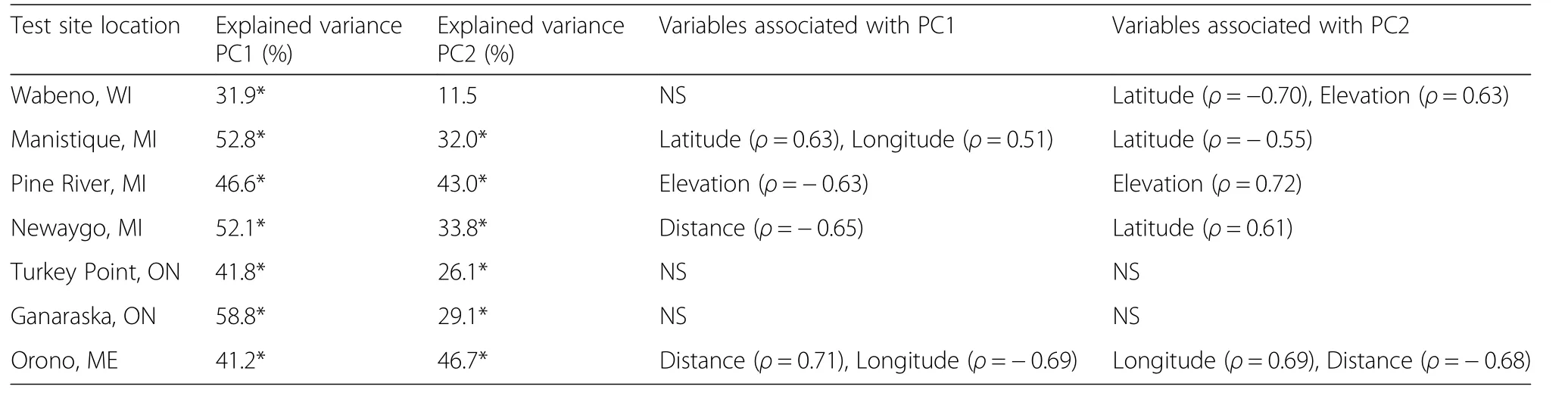
Table 4 Amount of explained variance for the principal component loadings of provenances at each test site on the principal component axes and correlation with geographical variables of provenance origin
Similar to mean maximum temperature,white pine radial growth responsesto minimum temperatureexhibited negative relationships of PC1 scores with the winter seasonal period[December(t-1)to February(t)]at Manistique and Newaygo,and Orono[January(t)to March(t)](Fig.4).At these sites,the dendroclimatic regression models generally had weaker explanatory power in connection with mean minimum temperature(as reflected by lower R2values)compared to the growth response to mean maximum temperature.The PC1 year scores at Turkey Point were negatively related to July(t)minimum temperature and PC1 year scores at Ganaraska were primarily negatively related with April(t-1)minimum temperature.The year scoreson PC2 indicated that radial growth at most test sites did not respond to mean minimum temperature except for positive responses primarily in the fall(t-1)at Manistique and negative responses to September(t)minimum temperature at Turkey Point(Fig.4).
Responses of radial growth to mean temperature reflected in the PC1 year scores showed similarities in responses to mean maximum temperature among test sites,except at Pine River where a secondary and positiveresponse to fall(t-1)mean temperature was evident(Fig.5).Furthermore,PC1 year scores for Orono were the reverse of rankings of the variables that were influential in the regression model,with more explanatory power provided by mean temperature in the winter seasonal period(t).The PC2 year scores responded positively to mean temperature in May(t)at Wabeno and fall(t-1)at Manistique(Fig.5).The PC2 year scores at Turkey Point were negatively related to mean temperature in June(t)at Newaygo,September(t)at Turkey Point,July(t)at Ganaraska,and August(t-1)at Orono.
The year scores of both PC1 and PC2 were significantly related to monthly and seasonal precipitation at most test sites(Fig.6).The PC1 year scores were mainly positively related with winter precipitation at Wabeno and Newaygo[December(t-1)],and Manistique and Turkey Point[December(t-1)to February(t)].In contrast,the PC1 year scores were negatively associated with May(t-1)precipitation at Ganaraska.The PC2 year scores were primarily and positively related with summer precipitation in July(t)at Wabeno,July(t)to September(t)at Newaygo,August(t)at Ganaraska,and July(t)to September(t)at Orono(Fig.6).
Year scores on both the PC1 and PC2 axes were significantly related to CMI at most of the test sites except for Ganaraska(Fig.7).The PC1 year scores were mainly positively related with winter CMI at Wabeno,Manistique,and Pine River[December(t-1)],and Turkey Point[December(t-1)to February(t)].The PC2 year scoreswere positively related with CMIin February(t)at Manistique and during the summer seasonal period(t)at Newaygo and Orono.In contrast,PC2 year scores were primarily negatively related to CMI in August(t-1)at Wabeno as well as the fall seasonal period(t-1)at Pine River and January(t)at Turkey Point.
Influence of location of provenance origin and bioclimate on radial growth response at each test site
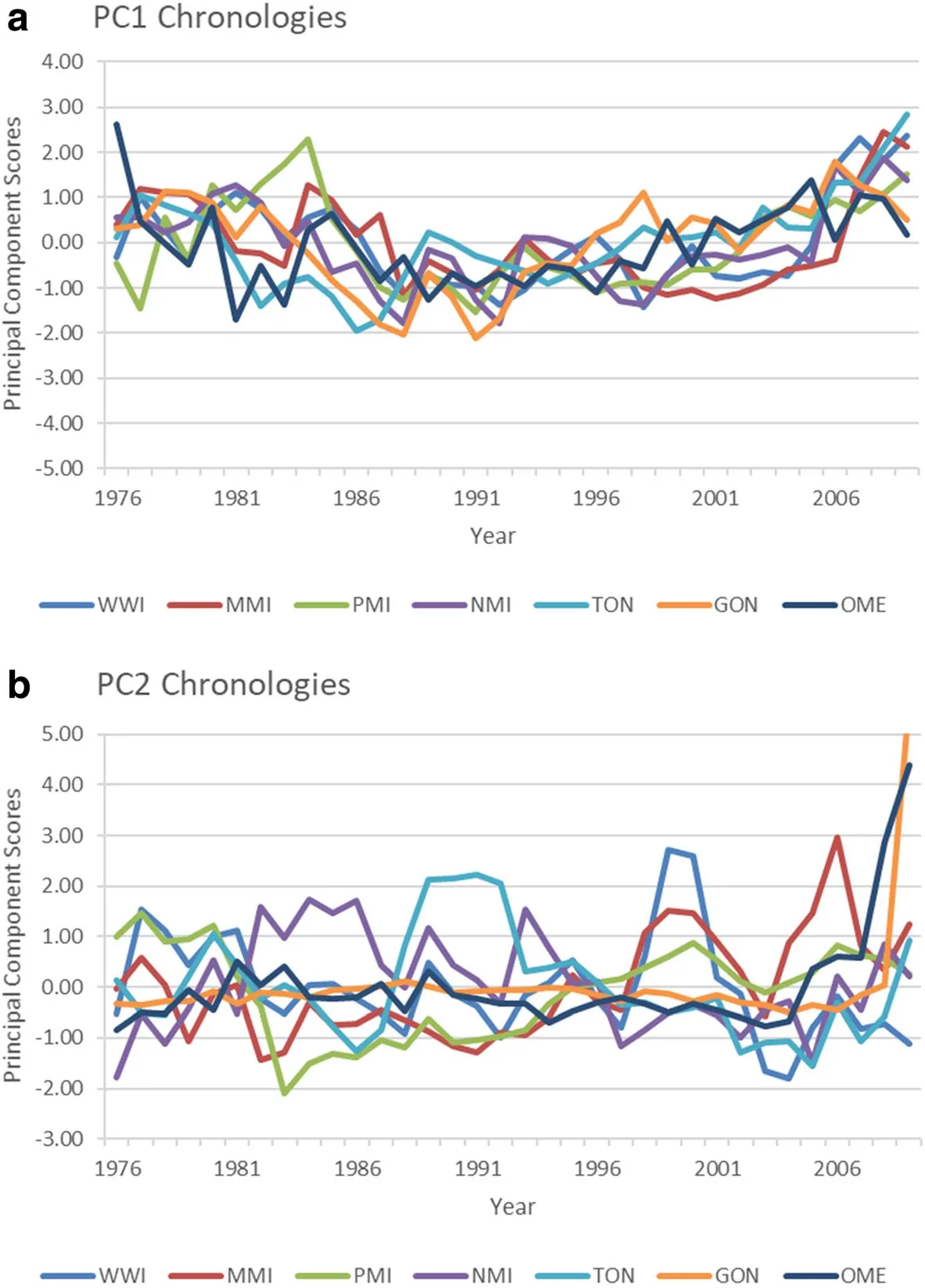
Fig.2 Time series chronology of the year scores of radial growth on the first principal component(PC1)(a)and second principal component(PC2)(b)axes at each of the 7 test sites.WWI=Wabeno,WI;MMI=Manistique,MI;PMI=Pine River,MI;NMI=Newaygo,MI;TON=Turkey Point,ON;GON=Ganaraska,ON;OME=Orono,ME
Significant correlations between radial growth responseand geographic and bioclimatic parameters of the provenances were also observed at the test sites.The loadings of the provenanceson PC1 werepositively correlated with latitude and longitude at Manistique,negatively correlated with elevation at Pine River,negatively correlated with distance from test site at Newaygo,and negatively correlated with longitude and positively correlated with distance from test site at Orono(Table 4).The loadings onto PC2 were negatively correlated with both latitude and elevation at Wabeno,negatively correlated with latitude at Manistique,positively correlated to elevation at Pine River,and positively correlated with longitude and negatively correlated with distance from test site at Orono(Table 4).The loadings of each provenance on PC1 and PC2 at each test site are provided in Table 5.
The loadings of the provenances on PC1 were not significantly correlated with the bioclimatic parameters of the location of provenance origin for the Wabeno and Orono test sites(Table 6).At Manistique,of the 7 significant relationships the provenance loadings on PC1 were significantly and most strongly correlated with precipitation of the driest quarter(+ρ)and temperature of the wettest quarter(?ρ).The two strongest correlations with the PC1 loadings at Pine River included temperature of the coldest quarter(?ρ)and precipitation of the wettest quarter(?ρ).The two strongest correlations with the PC1 loadings at Turkey Point were temperature annual range(?ρ)and mean diurnal range(+ρ).The provenance loadings onto PC1 at Ganaraska were significantly correlated with precipitation of the wettest quarter(?ρ)and temperature of the coldest quarter(?ρ).

Fig.3 White pine(Pinus strobus L.)radial growth represented by the year scores on the first and second principal component axes and the relationship with mean maximum temperature based on multiple regression analysis.Black boxes indicate a negative relationship and grey boxes indicate a positive relationship between growth and climate.Where more than one climatic variable was significant,numbers for a given test site indicate order of significance(1 is most important)based on standardized partial regression coefficients.All relationships shown are statistically significant(p<0.05).R2 is the adjusted coefficient of determination.NSindicates that no relationships were statistically significant
The loadings of the provenances onto PC2 at Wabeno were significantly correlated with 11 variables,with temperature of the warmest quarter(+ρ)and temperature of the wettest month(?ρ)having the strongest correlations(Table 7).A total of 7 significant correlations were present among the PC2 provenance loadings at Manistique in which the highest values were for the response to temperature of the wettest quarter(+ρ)and isothermality(+ρ).The PC2 loadings at Pine River were correlated with 4 parameters,with the correlation being highest with temperature of the coldest quarter(+ρ).The PC2 provenance loadings at Newaygo were significantly correlated with 6 biophysical parameters,with the 2 strongest correlations with isothermality(?ρ)and temperature of the wettest quarter(?ρ).The PC2 provenance loadings were correlated with 11 biophysical parametersat Turkey Point,with temperature annual range(+ρ)and temperature of the coldest month(?ρ)having the two strongest correlations.At Ganaraska,the PC2 provenance loadings were significantly correlated with 3 biophysical parameters,with the correlations with precipitation of the wettest period(+ρ)and temperature of the coldest quarter(+ρ)being the two strongest.PC2 loadings for Orono were not correlated with any bioclimatic parameter.
Discussion
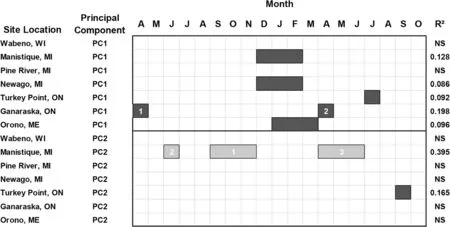
Fig.4 White pine(Pinus strobus L.)radial growth represented by the year scores on the first and second principal component axes and the relationship with mean minimum temperature based on multiple regression analysis.Legend descriptionsare provided in Fig.3 caption
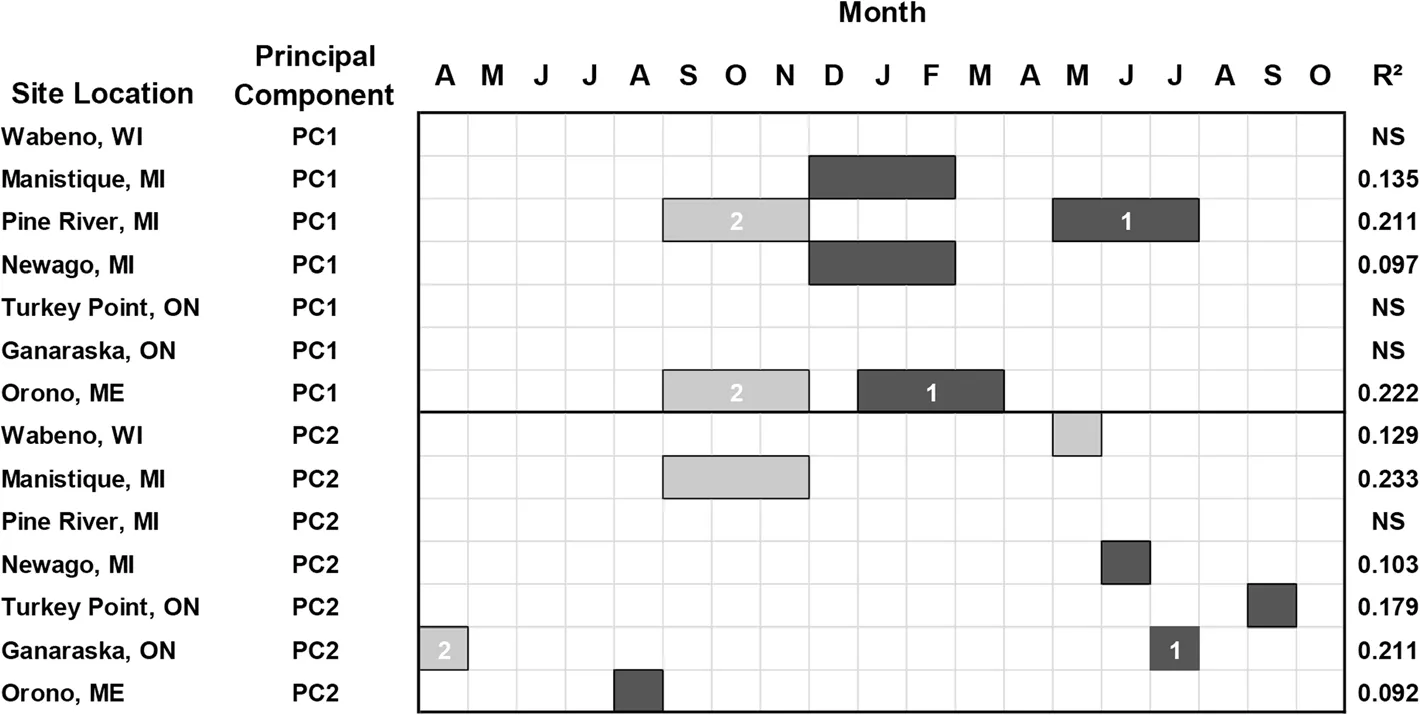
Fig.5 White pine(Pinus strobus L.)radial growth represented by the year scores on the first and second principal component axes and the relationship with mean temperature based on multiple regression analysis.Legend descriptions are provided in Fig.3 caption
Natural populations of white pine have been examined in prior dendroclimatic and dendroecological studies in eastern North America(Graumlich 1993;Abrams et al.2000;Black and Abrams 2005;Kilgore and Telewski 2004;Kipfmueller et al.2010,Chhin et al.2013).This study represents the first dendroclimatic assessment of a long-term range-wide provenance study of white pine,which consisted of 7 test sites in the United States and Canada and 13 provenances obtained from across the species’natural range.The results revealed complex radial growth responses to climatic and geographical variablesamong and within provenancesof whitepine.
Shared provenance radial growth responses to climate of each test site
Examination of year scores of the first and second principal component axes provided insight into similarities in climatic sensitivity shared among provenancesplanted at each of the 7 test sites.The negative relationships between the PC1 and PC2 year scores and temperature variables indicated that radial growth of white pine was likely inhibited by higher temperature,and thiswasobserved at all sitesexcept Wabeno(Figs.3,4 and 5).The radial growth response to high temperature also differed in seasonality.That is,a winter seasonal effect was evident at Manistique,Newaygo,and Orono;a spring effect at Ganaraska,and a summer effect at 5 of the 7 sites:Pine River,Newaygo,Turkey Point,Ganaraska,and Orono.From an ecophysiological perspective,high temperature stress can result in respiratory depletion of carbohydrate reserves that might otherwise be used to support radial growth(Fritts 1976;Kozlowski et al.1991).During the growing season,high temperature stress can also inducewater stressby increasing evaporativewater demand(Fritts 1976).High temperature has been observed to reduce radial growth of white pine growing in the southern region of the Lower Peninsula of Michigan(Chhin et al.2013)and in northern Minnesota(Kipfmueller et al.2010).During the winter,high temperatures can interfere with maintenance of cold hardiness leading to low temperature injury(Havranek and Tranquillini 1995).As well,periodic exposure to brief periods of warm temperatures followed by freeze events can result in leaf tissue injury and reduce the growth of coniferous tree species(Havranek and Tranquillini1995).
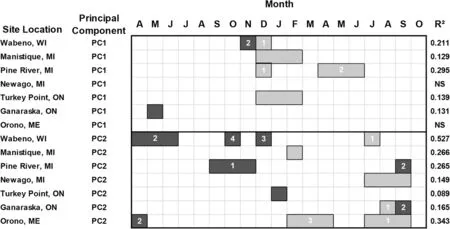
Fig.6 White pine(Pinus strobus L.)radial growth represented by the year scores on the first and second principal component axes and the relationship with mean total precipitation based on multiple regression analysis.Legend descriptionsare provided in Fig.3 caption
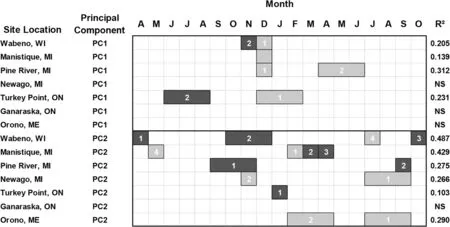
Fig.7 White pine(Pinus strobus L.)radial growth represented by the year scores on the first and second principal component axes and the relationship with the mean climatic moisture index based on multiple regression analysis.Legend descriptions are provided in Fig.3 caption
Another common relationship found between white pine radial growth(represented by the PC1 and PC2 year scores)and climate is that cold temperature stress either at the beginning(i.e.,spring)and(or)end of the growing season(i.e.,autumn)was correlated with reduced radial growth.This relationship was observed at 5 of the 7 sites:Wabeno,Manistique,Pine River,Ganaraska,and Orono.Conversely,the positive growth response to warmer spring and fall temperatures indicated that warmer conditions in the spring and(or)fall at these 5 test sites would favor radial growth by increasing the length of the growing season(Fritts 1976;Kozlowski et al.1991).The timing of the start of the growing season was found to be a factor affecting radial growth in a population of white pine growing in the northern region of the Lower Peninsula of Michigan(Kilgore and Telewski 2004),as well as Wisconsin and the Upper Peninsula Michigan(Graumlich 1993).In contrast,Chhin et al.(2013)examined a more southern population of white pine in Michigan’s Lower Peninsula and did not observe any radial growth dependency on spring temperatures.
White pine radial growth responses to precipitation and CMI varied with test site location.Generally,thePC1 year scores indicated that provenances of white pine at 4 of the test sites(i.e.,Wabeno,Manistique,Pine River,and Turkey Point)had a strong,positive response to winter precipitation and CMI.Positive responses to winter precipitation are usually indicative of increased snowpack that can help insulate the soil and prevent the development of cold soil temperatures causing root injury.Furthermore,higher snowfall accumulation increases ground water recharge during spring snow melt,providing abundant soil moisture to support radial growth early in the growing season (Fritts 1976;Kozlowski et al.1991).The PC2 year scores generally showed a positive response to summer precipitation at 4 of the 7 test sites:Wabeno,Newaygo,Ganaraska,and Orono.Radial growth was sensitive to summer drought stress at these test sites.Reduced radial growth due to growing season drought has been observed in white pine in other regions of eastern North America,including northern Minnesota(Kipfmueller et al.2010),the southern region of the Lower Peninsula of Michigan(Chhin et al.2013),an old growth stand of white pine in Massachusetts(Abrams et al.2000),and the southern Appalachian region(Vose and Swank 1994).
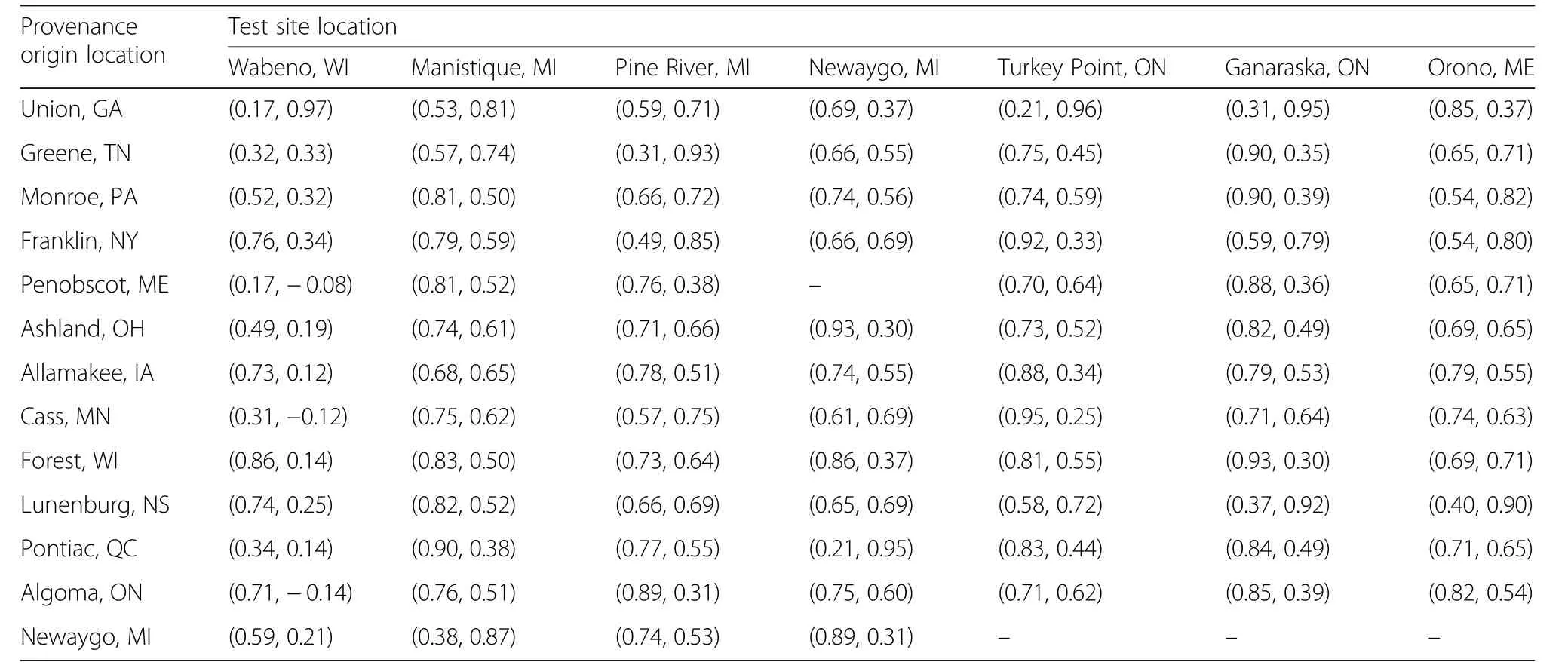
Table 5 Loadings of each provenance on the first(PC1)and second(PC2)principal component axis for each test site
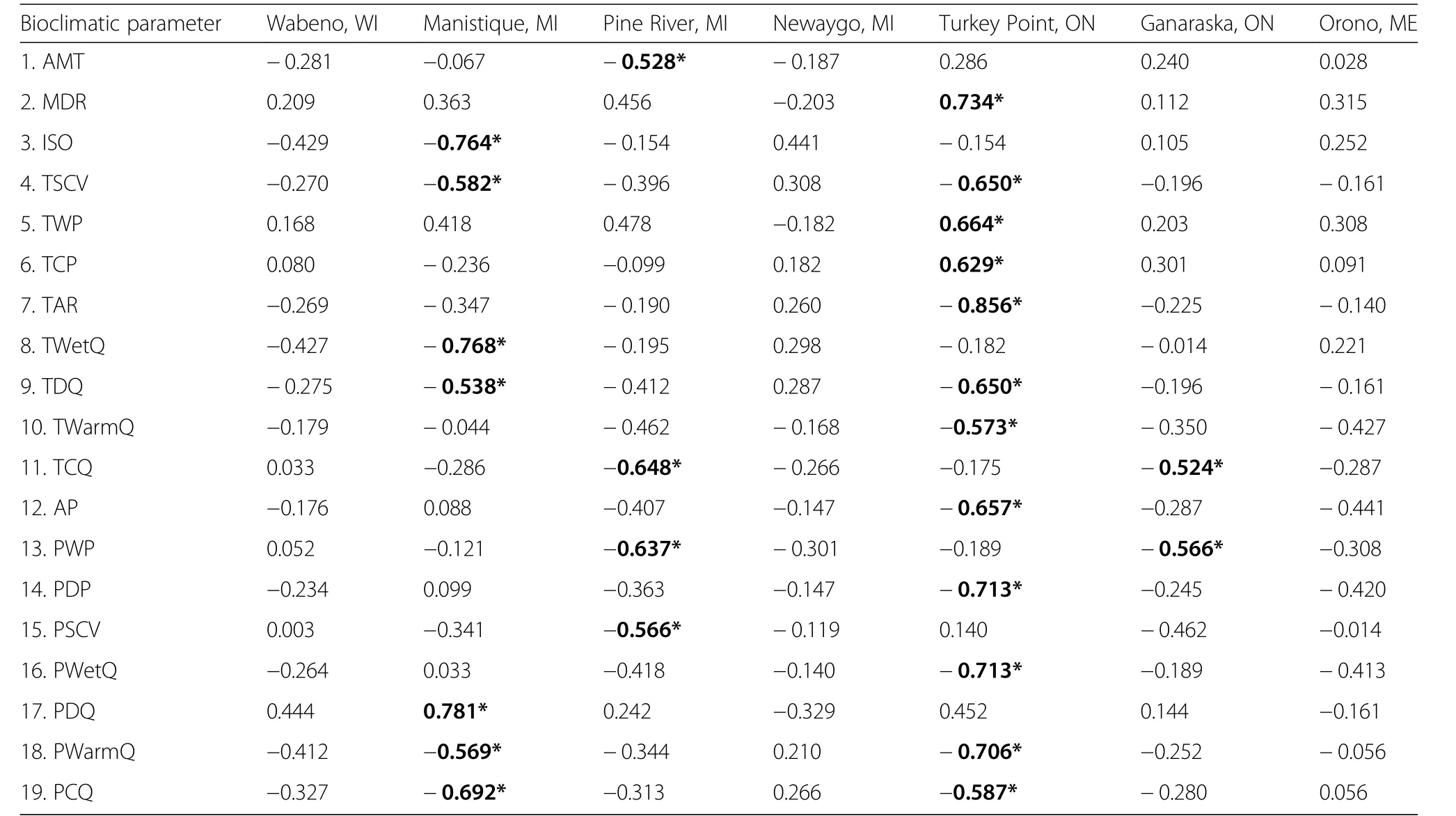
Table 6 Spearman rank correlation coefficients reflecting the sensitivity of the first principal component(PC1)loadings of the provenances at each test site with bioclimatic parameters of provenance origin location
The dendroclimatic regression models revealed significant negative relationships between white pine radial growth(i.e.,PC1 but mainly PC2 year scores)and spring and fall precipitation and(or)CMI at 5 of the 7 test sites:Wabeno,Manistique,Pine River,Ganaraska,and Orono.This relationship may be associated with storm damage to tree crowns resulting from exposure to high wind speeds(Everham and Brokaw 1996;Peterson 2000).Test sites near the Great Lakes region may have experienced more frequent wind storms during spring and fall(Scott and Huff 1996).Damage to tree crowns by windstorms could lead to reduced leaf area and decreased photosynthetic capacity.As well,subsequent carbohydrate allocation to crown repair would occur at the expense of reduced allocation to radial growth.At Wabeno and Turkey Point,the negative radial growth responses to winter precipitation could be the result of mechanical damage to crowns due to heavy winter snowfall.Winter crown damage may reduce growth the following growing season when trees prioritize crown repair at the expense of stem diameter growth(Havranek and Tranquillini 1995).
Influence of location of provenance origin and bioclimate on radial growth response at each test site
Examination of the loadings of the provenances on the first and second principal component axes provided an understanding of the primary geographic and bioclimatic parameters that contributed to the main separation of theindividual provenances at each test site(Tables 5,6 and 7).Separation of the loadings of the white pine provenances on the principle component axes at each site indicated that radial growth was affected by differences in the latitude,longitude,and elevation of provenance origin,depending on test site location.This provides evidence for the existence of genetic variation of radial growth potential among white pine provenances,likely associated with spatial variation in the temperature and precipitation regime across its natural range.These resultsare consistent with earlier findings for this trial(Wright et al.1970)and the results of other white pine provenance tests(Genys 1987).In contrast,results from a provenance study of European beech(Fagussylvatica L.)indicated that variation in radial growth response to temperature of northerly and southerly provenances were similar and primarily under environmental control,with limited differences due to genetic background of the provenances(Eilmann et al.2014).In another study,high gene flow limited local genetic adaptation resulting in similarities in growth response to climate in both European larch(Larix decidua Mill.)and Norway spruce(Picea abies(L.)Karst.)provenances along an elevational gradient.
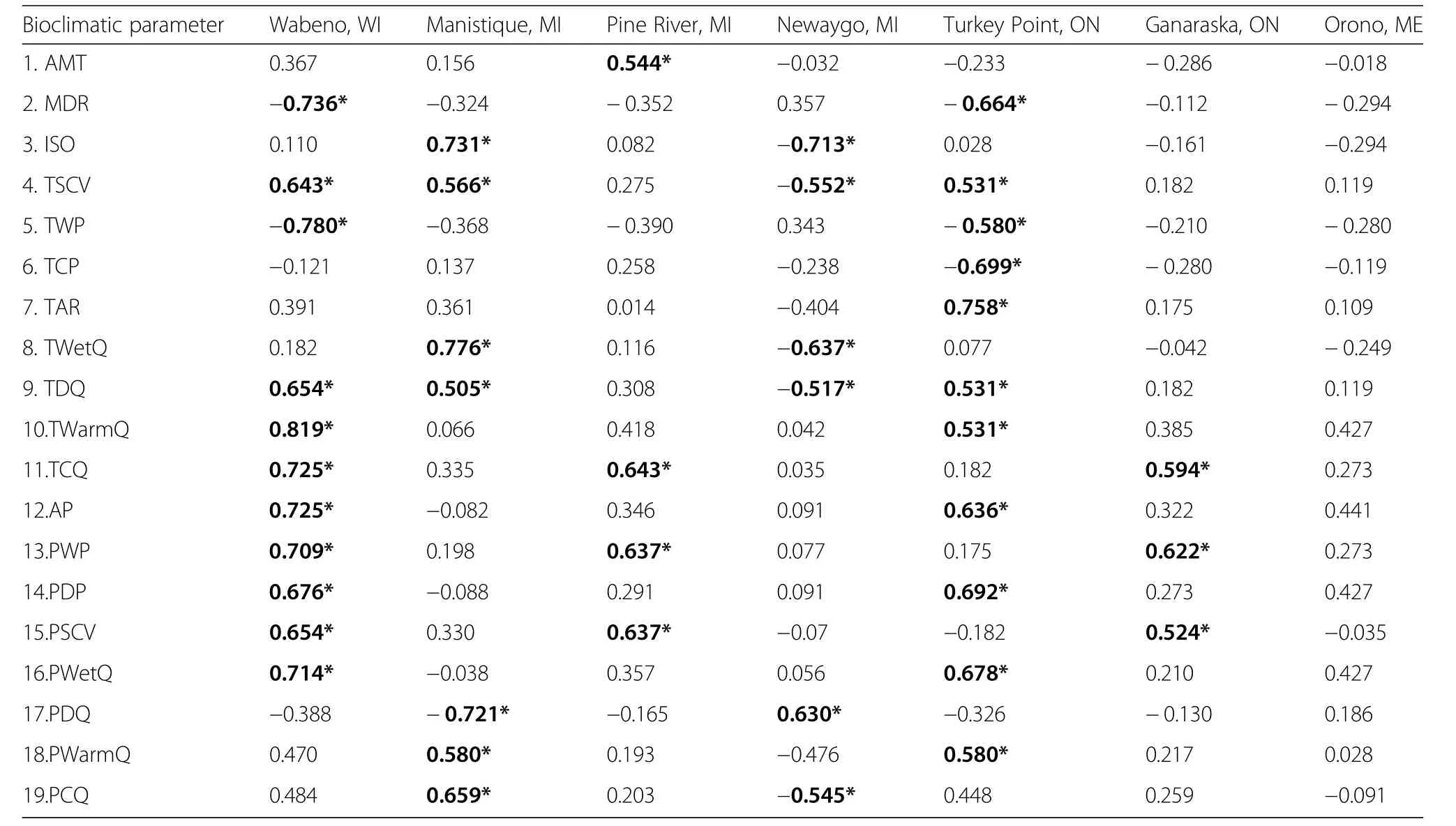
Table 7 Spearman rank correlation coefficients reflecting the sensitivity of the second principal component(PC2)loadings of the provenances at each test site with bioclimatic parameters of provenance origin location
Responsiveness of provenance loadings to bioclimatic parameters also varied with test site.Separation of provenance loadings on the PC1 axis was significantly correlated with bioclimatic parameters at 4 of the 7 sites:Manistique,Pine River,Turkey Point,and Ganaraska.Of these 4 sites,temperature-related bioclimatic parametershad thestrongest correlation with the PC1 provenance loadings,while a precipitation-related bioclimatic parameter was significant at Ganaraska.Furthermore,Turkey Point had the most bioclimatic parameters correlated with the PC1 provenance loadings.All test sites,except for Orono,had significant correlations among the PC2 provenance loadings.Both Wabeno and Turkey Point had the most bioclimatic parametersassociated with the PC2 provenance loadings.Overall,to some degree,bioclimatic parameters were significantly associated with provenance loadings and hence differences in radial growth responses at most of the test sites,except for Orono.Of these 6 test sites,5 were primarily affected by temperature-related bioclimatic parameters while Ganaraska was driven more by a precipitation-related bioclimatic parameter.This result conforms with the general understanding that temperature is the most important driver of adaptive genetic variation in growth,shoot phenology,survival,and frost hardiness of conifers(Gray et al.2016).Our results also suggest that at the Orono test site,differences in radial growth were likely not affected by bioclimatic parameters of the provenances,and based on dendroclimatic analysis of the year scores,the provenances had similar responses to climatic conditions.Most dendrochronological studies of genetic variation in tree species have examined multiple provenancesat a singletest site(e.g.,Eillman et al.2014),but our study is one of only a few that have examined radial growth sensitivity of multiple provenances established at multiple test sites.A combination of both site and provenance related sensitivities of radial growth to climate were reported for lodgepole pine(Pinuscontorta Dougl.var.latifolia Engelm.)in western Canada,with provenances with warm climates grown at warm sites responding mainly to summer dryness,while provenances from cold climates planted at warm site were generally responsive to temperature(McLaneet al.2011).
The information presented in our study could be used to help identify climate adapted provenances of white pine for planting in the northern portion of its natural range.For example,focusing only on the component loadingsof PC1 and the two highest observed correlations,our results suggest that the best radial growth in the current climate of Manistique might be achieved by planting white pine provenances with comparatively high precipitation in the driest quarter,and relatively lower temperature during the wettest quarter.Aswell,the absence of significant correlationsfor the Orono test site suggests that white pine may be generally well adapted to the maritime climate of this region,regardless of provenance.Clearly,a more detailed,comprehensive assessment of genetic variation in white pine is necessary to make confident provenance selections to support assisted migration efforts to better adapt forests to climate change(Yang et al.2015).However,our results suggest that broad scale genetic variation in white pine radial growth response does exist across a large portion of its natural range,and this variation is expressed when provenances are planted outside their regional climatic habitat.
Conclusions
The future regional distribution,abundance,and productivity of white pine and white pine forest types under a changing climate have been projected using a variety of modelling approaches and climate change scenarios(McKenney et al.2007;Joyce and Rehfeldt 2013;Peters et al.2013;Handler et al.2014;Iverson et al.2008,2017),each having a different structure,limitations,assumptions,and levels of uncertainty(Iverson et al.2017).Bioclimatic,or species distribution models,have typically shown the area of suitable climatic habitat for white pine to decrease and/or move northward by the end of the century,with larger changes under higher greenhouse gas emissions scenarios,but the degree of change varies with region(McKenney et al.2007;Joyce and Rehfledt et al.2013;Iverson et al.2008,2017).For example,the future abundance of white pine in three forest regions of the United States was unchanged,or experienced small to large reductions by the end of the century,depending on the forest region and the model used,with projected changes being larger in scenarios where future greenhouse gas emissions remain comparatively high and the climate changes more rapidly(Iverson et al.2017).In the northern portion of its natural range,white pine may experience modest increases in growth during this century under a slightly warmer climate(Peters et al.2013;Handler et al.2014;Yang et al.2015).Conversely,white pine growing near its warmer range limit,may be poorly adapted to increasing temperatures and suffer significant reductions in growth and survival under climate change(Joyce and Rehfeldt 2013;Yang et al.2015).
Future climate change presents both challenges and opportunities for management of white pine.The current study indicates potential vulnerabilities of white pine radial growth in the northern portion of its range to higher temperatures,summer moisture stress,and crown damage by high winds and heavy snowfall.Projected future increases in temperature,summer drought stress,and higher frequencies of storm activity(IPCC 2007)in this region will require efforts to both adapt management practices to the future climate and enhance conservation of genetic resources of white pine.On the other hand,the current study suggests that white pine radial growth at some test site locations has been historically limited by cold spring and fall conditions.Since future climatic warming will likely increase the length of the growing season,increased growing season temperature could potentially offset these growth limitations in colder regions of its natural range.These results are consistent with those of Yang et al.(2015)that reported optimal height growth of young white pine occurs at an AMT of about 11°C,and that white pine in relatively colder locations may benefit,at least initially,from modest climate warming.
Additional file
Additional file 1:Chronology statistics(i.e.,interseries correlation coefficient and mean sensitivity)for each seed source at each test site location.(XLSX 16 kb)
Acknowledgements
Thanks go to N.Eskelin,B.Brown,J.Winters,P.Lu,S.Colombo,K.Finley,K.Minnix,I.Allen,B.Birr,and A.Wiese for assistance with field and laboratory data collection.We are grateful to E.Bauer,Dr.W.Headlee,and two anonymous reviewers for their many helpful comments on earlier versions of this manuscript.The conclusions and opinions in this paper are those of the authors and not the Northeastern States Research Cooperative(NSRC),the USForest Service,or the USDA.
Funding
This study was supported by the Northeastern States Research Cooperative(NSRC)through funding made available by the USDA Forest Service.
Authors’contributions
RZ,WP,JBdesigned the field sampling and collected the data;SCanalyzed the data;SClead the manuscript writing with contributions from RZ,WP,and JB.All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Author details
1Division of Forestry and Natural Resources,West Virginia University,322 Percival Hall,POBox 6125,Morgantown,WV 26506,USA.2USDAForest Service,Northern Research Station,5985 Highway K,Rhinelander,WI54501,USA.3Ontario Forest Research Institute,1235 Queen St.E,Sault Ste.Marie,ONP6A 2E5,Canada.4USDAForest Service,Northern Research Station,271 Mast Road,Durham,NH 03824,USA.
Received:6 November 2017 Accepted:13 February 2018

- Forest Ecosystems的其它文章
- Predicting the provisioning potential of forest ecosystem servicesusing airborne laser scanning data and forest resource maps
- Detecting treeline dynamics in response to climate warming using forest stand maps and Landsat data in a temperate forest
- Nutrient retention and release in coarse woody debris of three important central European tree species and the use of NIRS to determine deadwood chemical properties
- Species-specific,pan-European diameter increment models based on data of 2.3 million trees
- Drought can favour the growth of small in relation to tall trees in mature stands of Norway spruce and European beech
- Forests,atmospheric water and an uncertain future:the new biology of the global water cycle