
Neuroanatomy and morphological diversity of brain cells from adult crayfish Cherax quadricarinatus*
2018-12-22 07:00:44DUANHu段虎JINSongjun金松君LIFuhua李富花ZHANGXiaojun張曉軍XIANGJianhai相建海
關(guān)鍵詞:張曉軍
DUAN Hu (段虎), JIN Songjun (金松君) , LI Fuhua (李富花) , ,ZHANG Xiaojun (張曉軍), , XIANG Jianhai (相建海) , ,
1 Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071,China
2 Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China
Abstract As in vertebrates, brains play key roles in rhythmic regulation, neuronal maintenance,differentiation and function, and control of the release of hormones in arthropods. But the structure and functional domains of the brain are still not very clear in crustaceans. In the present study, we reveal the structural details of the brain in the redclaw crayfish using hematoxylin-eosin staining and microscopic examination, firstly. The brain of crayfish is consist of three main parts, namely, protocerebrum,deutocerebrum, and tritocerebrum, including some tracts and commissures, briefly. Secondly, at least 9 kinds of brain cells were identified on the basis of topology and cell shapes, as well as antibody labeling. We also provide morphological details of most cell types, which were previously un-described. In general, four types of glia and three types of neurosecretory cells were described except cluster 9/11 and cluster 10 cells.Glia were categorized into another three main kinds: (1) surface glia; (2) cortex glia; and (3) neuropile glia in addition to astrocytes identified by GFAP labelling. And neurosecretory cells were categorized into I, Ⅱand III types based on morphological observation. Finally, cluster 9/11 and 10 cells derived from the brain of crayfish, could be used for primary culture about 7–9 d under the optimized conditions. There results provide a resource for improving the knowledge of the still incompletely defined neuroendocrinology of this species. Using the crayfish as an animal model, we are easy to carry out further research in manipulating their endocrine system, exploring cellular and synaptic mechanisms so much as larval production on a small scale, such as in a cell or tissue.
Keyword: Cherax quadricarinatus; brain’s structure; brain cells; primary culture
1 INTROD UCTION
The brain (cerebral ganglia) of crayfish could serve as a good experimental model in arthropods. Because aparting from their importance as known keystone species in the food web of freshwater ecosystems,redclaw crayfish have many notable virtues, such as large size, simple nervous system with well-defined neural circuits, and behaver in very interesting patterns,fielding from reflexes to complicated social interactions and so on (Lindsey and Tropepe, 2006). The brain of crustacean is located in the head at the anterior end, at up to the esophagus and also called for supraesophageal ganglion (Sandeman et al., 1992; Smarandache-Wellmann, 2016). Initial studies on the brain of spiny lobster in 1960’s (Maynard, 1962) were untimely terminated. Before 1990’s, the endocrine systems in eyestalks of crayfish were investigated (Cooke and Sullivan, 1982; álvarez et al., 2005; Manfrin et al.,2015). Since 2000’s, the cerebral ganglion of crustaceans has been gained renewed research interest for replenishment of neural progenitors (Hartenstein,2014), extensive neuromodulation by a diverse set of signaling peptides (Ye et al., 2015), and control of sexual maturation in adult crayfish by various neurohormones (Sullivan and Beltz, 2005; Lindsey and Tropepe, 2006; Stegner et al., 2015).
Adult neurogenesis have been well certified in the brains of vertebrates, such as in mammals, fish,amphibians, reptilesand birds, which are regarded as a constitutive procedure in the brains of many arthropods (Lindsey and Tropepe, 2006; Schmidt and Derby, 2011). In the brains of some insect species,adult neurogenesis generated new local interneurons(Kenyon cells) of the mushroom bodies (Ghosal et al.,2009; Fernández-Hernández et al., 2013; Ito et al.,2014). However, limited information were regarding the biological function identification for the new generation neurons in crustaceans brains (Sullivan and Beltz, 2005). Similar to most mammals and insects, life-long neurogenesis was a notable feature of the brains of Crustacea, Decapoda (Benton and Beltz, 2002; Hansen and Schmidt, 2004). Most regions of the decapod brain ceased to exhibit neuronal proliferation when embryonic precursor cells disappeared, except for the central olfactory lobe, where mitotic activity continueed throughout life (Benton and Beltz, 2002; Beltz and Sandeman,2003). According to recent research, hemocytes could be an origin of adult-born neurons inProcambarus clarkii, and suggested that the immune cells played an important contributor to the neurogenesis of the adult crayfish (Benton et al., 2014). Neuronal proliferation had been reported mainly three different cells from different zones or parts related to the olfactory pathway; however, specific clusters, in which neurogenesis occured, varied among taxa (Schmidt and Harzsch, 1999; Sullivan and Beltz, 2005) in the brains of adult decapods.
The cerebral ganglion of decapods has gained increased research attention not only for its rhythmic regulation and neuronal maintenance, differentiation,and function, but also for its coordination, synthesis,and release of important hormones for controlling sexual maturation (Christie, 2011; Apitz and Salecker,2014). For example, gonadotropin-releasing hormone(GnRH) stimulated gonad maturation (Saetan et al.,2013) and synthesis of molt inhibiting hormones as well as maintained the integrity of the male genital system and promoted serotonin (5-HT) production in the central nervous system (CNS) of crustaceans(Saver et al., 1999). Moreover, dopamine (DA) in the brain were reported to inhibit testicular maturation inP.clarkiiand ovarian maturation inMacrobrachium rosenbergii(Tinikul et al., 2011). Furthermore,cerebral ganglion might play an important role in regulating the osmolarity of body fluids in the mud crab (Xu et al., 2010). Numerous peptide families and amines have been identified in crustacean brain through mass spectrometry and other methods(Christie, 2011; Ye et al., 2015; Zhang et al., 2015).Analysis results could be used in future neuropeptidome discovery studies on crustacean species without requiring prerequisite information regarding preprohormones (Ye et al., 2015) and developing specific antibodies. However, the precise locations of neuronal projections for a related omics study are quite difficult to illuminate because of lack of determined neuropil boundaries and specific antibodies. Defining the functional and chemical phenotypes of adult-generated cells is an important first step toward understanding the significance of persistent neurogenesis in the corresponding species.
Redclaw crayfish displays a number of physical,biological, and commercial attributes, which render the species as a suitable aquaculture candidate(Ghanawi and Saoud, 2012). Knowledge on the histology and classification of cells in brain is essential to identify specific neuron cells. The present study aims to describe the brain structure and cell types of the adult redclaw crayfish, determine suitable cell types for culture, and establish an approach to maintain neuronal growth in vitro. This study also proposes ganglionic and primary cell culture techniques for developing a useful model system in crustaceans to manipulate their endocrine system, exploring cellular and synaptic mechanisms so much as larval production and sexual maturation on a small scale.
2 MATERIAL AND METHOD
2.1 Animals
The specimens ofCheraxquadricarinatuswere collected from a commercial farm of Xiamen in Fujian Province, China. Adult male and female crayfish were maintained in a flow-through fresh water aquarium in the IOCAS (Institute of Oceanology,Chinese Academy of Sciences) laboratory at 24°C to 26°C with a light/dark cycle 12 h:12 h. The animals were acclimated for at least 1 week and fed with chopped mussels once a day. Inter-molt, healthy, and at least five adult crayfish (carapace lengths about 10–15 cm) were used for experiments. All the research were conducted in accordance with the Ethical Reference Frame for Biomedical Investigations of the Conicet equivalent to the standard procedures for animal care and use of the NIH of the USA. All efforts were made to minimize the number of animals used.
2.2 Brain isolation and tissue preparation
Crayfish were kept in the refrigerator about 20 min for anesthetization, brains were carefully dissected out crayfish in pre-cooled crayfish saline (Jiravanichpaisal et al., 2006) by removing the overlying carapace and connective tissues and then fixed in Davidson’s fixative for 12 h in darkness (Jemaa et al., 2014) at 4°C for Hematoxylin and eosin (HE) staining. Isolation was manipulated rapidly into the base of each eyestalk by using an ophthalmic forceps to wipe out nerve sheath covering the brain under stereomicroscope, maintaining the structural integrity of the brain, and preventing neuronal injuries. All brains of crayfish were dissected not in a specific time in addition to the night.
2.3 Histology and imaging
Tissue samples were dehydrated in 70% to 100%successive ethanol baths and then in xylene three times before embedding in paraffin (Sigma) (Jemaa et al., 2014). Cross-sections of 5–7 μm thick were cut using a microtome (Leica) and dried in an oven at 40°C for 2 h. Paraffin was eliminated in xylene bath twice, and the sections were rehydrated in successive 100% to 0 ethanol baths.
For histological analysis, tissue slides were incubated with hematoxylin for 3 min, counterstained for 0.5 min with eosin G, and washed twice in 100%ethanol and xylene before mounting. Pictures were obtained using a NIKON inverted microscope(NIKON TS100) and optimized for brightness and contrast by using Adobe Photoshop.
2.4 Immunohistochemistry
Brains were fixed in 4% paraformaldehyde and sliced as Section 2.3 described (Kress et al., 2016).Immunohistochemistry were done as described with some modification according to Histostain-Plus Kit.Briefly, after being blocked in 3% BSA, the paraffin sections with 5–8 μm thickness were first incubated with primary antibodies of GFAP (Proteintech, diluted at 1:100 with PBS) at 4°C for overnight. And then incubated with Alexa Fluor 488-labeled goat-antirabbit antibody (Abclonal, diluted at 1:100 with PBS)as the second antibody at room temperature for 60 min. Samples were washed three times with PBST,and incubated with 500 mL Hoechst33342 (Thermo Fisher scientific, diluted 1:5 000 in CPBS) and phalloidine (Sigma) about 60 min to stain the nucleus and cytoskeleton, respectively. Controls were carried out without primary antibody. Another five times of washing, samples were mounted in buffered glycerin(Solarbio) and observed by Laser Scan Confocal Microscope (Zeiss).
2.5 Single cell isolation and primary cell culture
Papain (Thermo Fisher Scientific) is a tissuespecific dissociation reagent for isolation of primary cortical and hippocampal neurons from the brain tissue. The tissues were treated with 2 mg/mL papain in crayfish phosphate buffer saline (CPBS) solution for about 30 min at 23±2℃. After dissociation, the tissue samples were rinsed 3 times to reduce effect of enzyme with precooled saline at 4°C, followed by a clean pipet for approximately 20 times to make the cell suspension become turbid. For light microscopy,100 μL of the cell suspension was placed on a clean cell culture dish. Accordingly, cells were categorized based on morphological observation, including cell size and shape, and examined through a NIKON TS100 microscope.
The dissociated cells from brains were added to small amounts of DNA enzyme. The cells were resuspended in optimized L-15 medium, and seeded into 24-well plates (Corning). After attachment for 1 h at room temperature to make sure cells for attaching to the bottom of plate, the culture medium was changed for the first time by adding to 100 U/mL penicillin,0.1 mg/mL streptomycin, and 2 mmol/L glutamine. The cells were maintained in a cell incubator at 25℃. The medium was replaced once every 2 d. The cells were examined daily with a NIKON inverted microscope.
3 RESULT
3.1 Structure of the brain (cerebral ganglia)
Different regions of brains were given names that facilitate descriptions of neural circuits, projections of specific neurons, and distribution of molecules associated with neural functions (Ito et al., 2014). The size of adult redclaw crayfish brain (except optic ganglia) was s approximate 1.0±0.2 mm in length and 1.6±0.3 mm in width, with carapace length of 10 to 15 cm. Similar to vertebrates and insects, the brain of crayfish exhibited a bilateral symmetry. A brain was divided into three main parts from superior to inferior,protocerebrum, deutocerebrum, and tritocerebrum,namely (Fig.1).

Fig.1 Graphic section of cell clusters and neuropile compartments in the ventral and dorsal views of the brain from redclaw crayfish
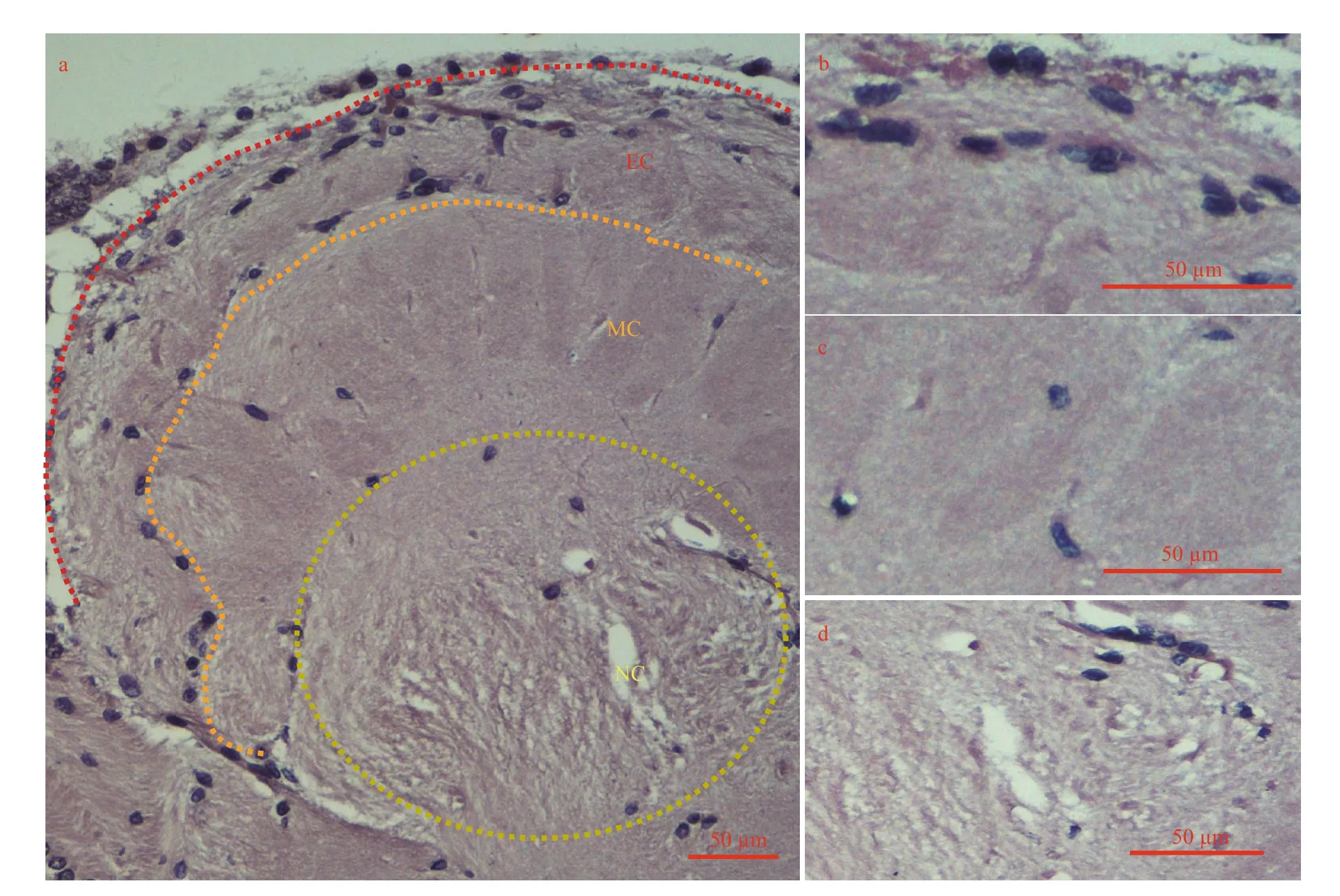
Fig.2 Three of the four types glia of distribution in the accessory lobe (AL) from the adult crayfish brain (horizontal section)
The protocerebrum was subdivided into the anterior (AMPN) and posterior medial protocerebrum(PMPN) connected by protocerebral tract (PT) in the middle of the sections. OG (optic ganglia) (Fig.1a) is composed of the periphery of lamina ganglionaris,medulla externa (ME), medulla interna (MI), medulla terminalis (MT), and sinus gland (another paper), not include in the protocerebrum. The posterior protocerebrum was composed of AMPN, PMPN, PB(Fig.1b, c, d) and the central body neuropil (CB).
As reference to previous literature (Sandeman et al., 1992; Zhang et al., 2009), deutocerebrum were mainly composed of OL, AL, LAN, and MAN. The OL lied to the front of AL (Fig.1b, c, e), presumably transfer chemoreceptors into antenna, but the AL seemingly had no chance direct input from primary axons. Moreover, the OL was innervated by some neurons, whose somata existed in 2/3 cell clusters,namely cluster 9/11 and 10 (Fig.1b, c, e). Both OL and AL were linked by some cells (OGTN), whose axons took shape the olfactory globular tract (OGT)(Fig.1d). Cell bodies associated with the olfactory and accessory lobes, cluster 9/11 and 10 were small with large nuclei (Fig.1c, e). According to previous reports, the LAN might receive afferent signals from mechanoreceptors, statocysts, and non-aesthetasc chemoreceptors from ipsilateral antenna (Fig.1b, c).Limited information was known about projections to the MAN of the brain.
The tritocerebrum was composed of antenna 2 neuropil (AnN), tegumental neuropils (TN),esophageal connective tracts (Ec) and commissures(Fig.1a, b, c, e). All of these neuropils lied posteriorly to the lateral antenna 1 neuropil and the accessory lobe and was visible in the middle of the section stacks. Medially, AnN was bent by approximately 90°to posterior and extended toward the Ec. Signal transmission or cell migration was mostly located in tracts. Neurosecretory cells were ocated at the last posterior-middle dorsal region of the brain. They were a cluster of large neurons, projecting fibers to the MAN (Fig.1b, c).
Insect glia represents different cell population and plays an important part in progenitor neurons and neurological function (Hartenstein, 2011). But almost nothing was known about how many types and what functionality for glia of crayfish. Four kinds of glia were distinguished in the brain. The AL area was composed of three layers (EC, MC, and NC) in the brain (Fig.2). The EC was surrounded by two layers of glial cells with circles in the outer layer of perineurial glia (surface glia) and an inner layer of sub-perineurial glia. The outer layer covered the entire brain. Nerve sheath was removed along with perineurial glia to collect an intact and clear brain.The MC was composed of cortex glia, and cells were mostly confined to the accessory glomeruli. Only a few clusters of cell on the NC were reticular. Glial cells were distributed in OL with scattering, except for exterior cortex (Fig.1e). Compaired with control(figure was not shown), astrocytes were located mainly in nerve sheath around the brain, they had a small amount of distributed between OL and AL(Fig.3).
3.2 Neuronal morphology
The whole brain was dissociated with Papain(Roche), of the 6 major cells were identified based on morphological diversity (Fig.4). Intact OL and AL were dissected away from the whole brain, and buffeted to make them become single cell as far as possible, other components were deal with the same method to harvest single cells. Five cells were analysed for each type of cells. Putative glia cells(Fig.4a–c) mainly existed in the OL and AL. Surface glia (Fig.4a) was roughly 22.5±2.5 μm (n=20) in diameter, with two dendrites (about 30 μm) from EC.Medial cortex glia (Fig.4b) was about 18.8±1.8 μm(n=20) in diameter with more than two dendrites(about 20 μm) from MC, and neuropile glia (Fig.4c)was about 18.4±1.7 μm (n=6) in diameter with two long dendrites greater than 50 μm might come from NC, mainly. Neurosecretory cells I–III might be derived from the central domain of the brain because these cells were not found in other regions, such as AL, OL, and clusters. Neurosecretory cell I (Fig.4d)was 23.6±2.1 μm (n=6) in minor axis and 43.2±8.1 μm(n=5) in macro-axis with abundant soluble substance,neurosecretory cell II (Fig.4e) was the largest of all types, with soluble substance of roughly 41.7±9.2 μm(n=7) in diameter without any dendrite or axons, and neurosecretory cell III (Fig.4f) taking on clusters, was 35.4±10.8 μm (n=7) in diameter with dendrite and axons.
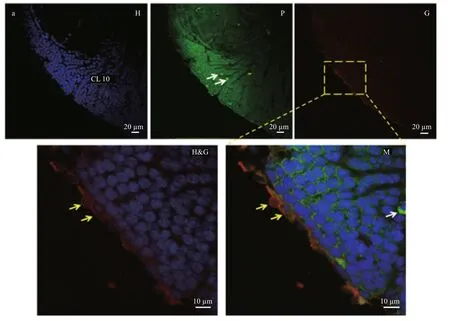
Fig.3 Distribution of astrocytes in the adult crayfish brain (horizontal section)
Fig.3 Distribution of astrocytes in the adult crayfish brain (horizontal section)
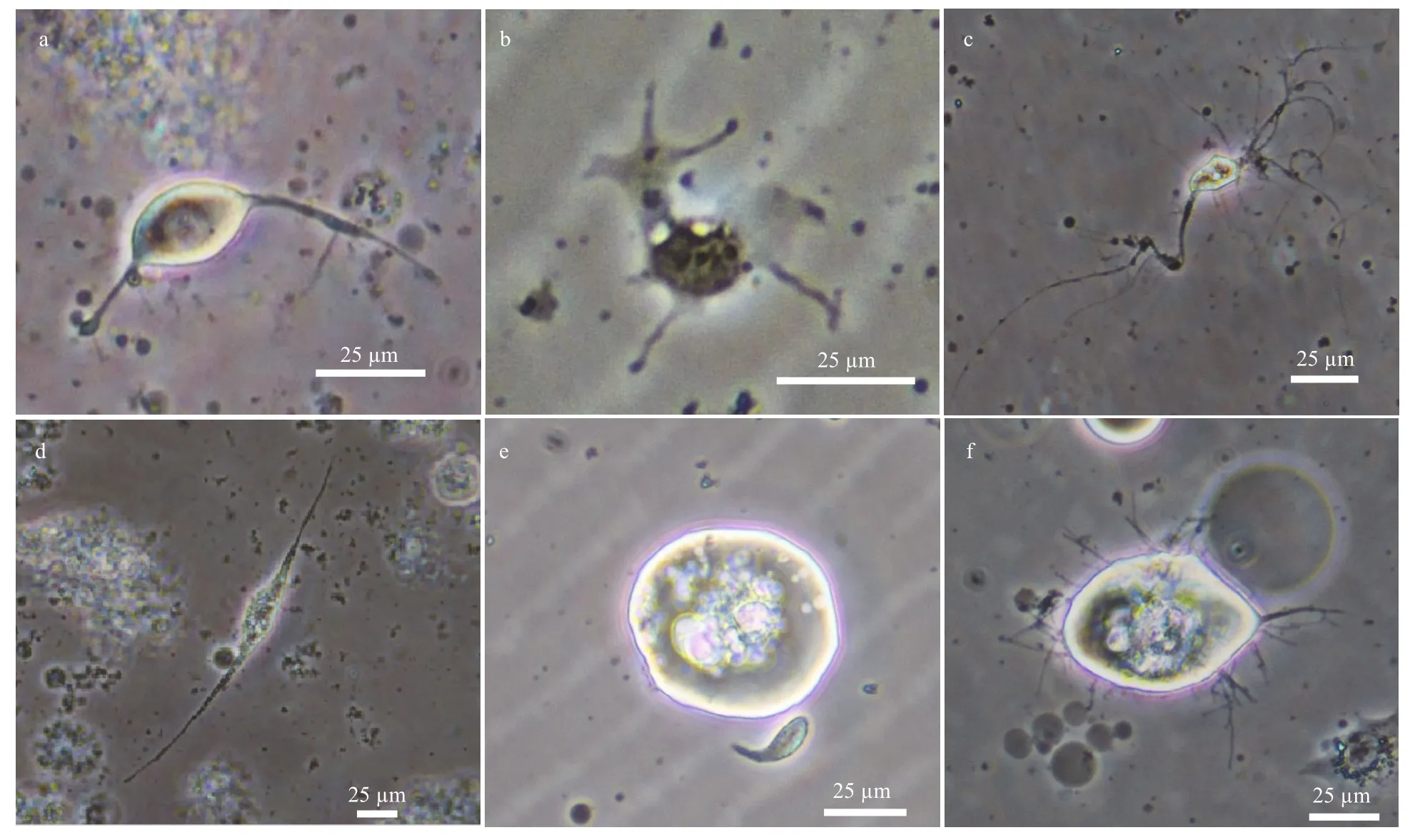
Fig.4 Morphological types of brain cells
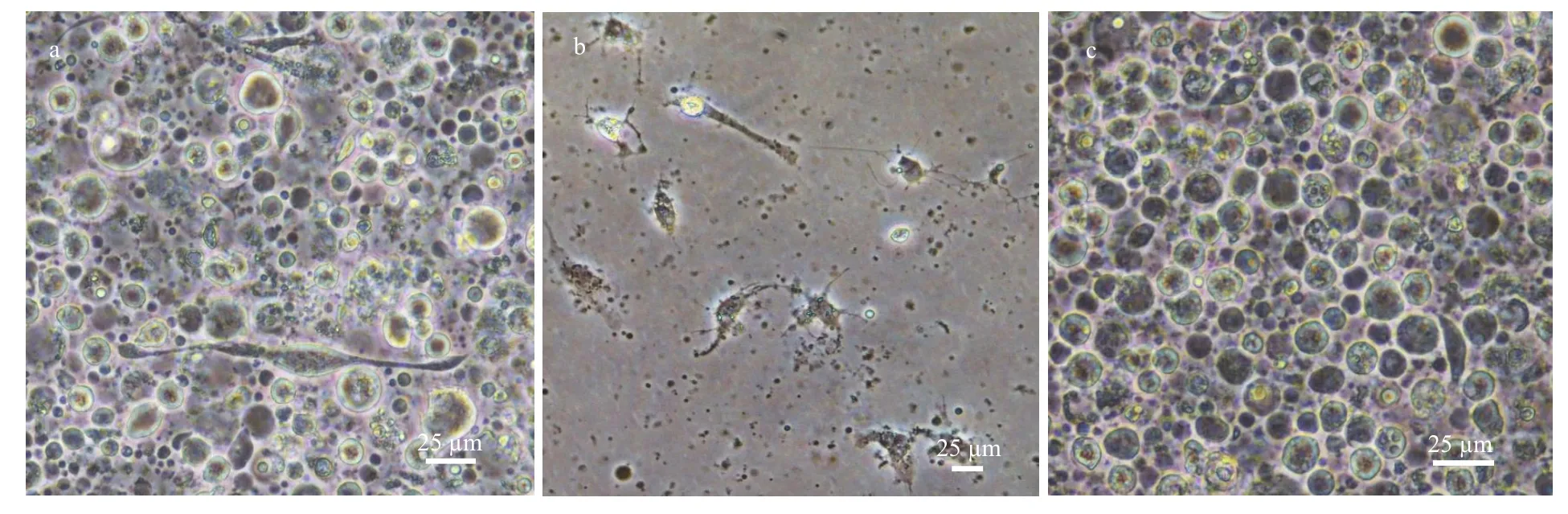
Fig.5 Cell cultures from different brain parts
3.3 Primary cell culture
Modified Liebowit’z L-15(ML-15) medium and Grace’s medium (Gibico) were used to examined which kind was suitable for growth of single cells from the redclaw crayfish neurons. Neurons from the crayfish brain were examined under inverted microscope and micro-photographed daily for consecutively until the cells lost typical morphology of neurons and diminished eventually. The results showed that ML-15 medium was more suitable for dissociated cells (figures are not shown). For whole brain cells,neurons could only survive 2 d or so (Fig.5a) but then retracted and formed debris. Glia cells (Fig.5b) from OL and AL could survive 5 d or so, and cells (Fig.5c)from CL9/11 and CL10 also could survive 5 d at least,mainly. It was no found that cells occurred in obviously proliferation from CL9/11 and CL10, though studies have been shown that cells could be labeled by BrdU from both clusters (Benton et al., 2013).
4 DISCUSSION
The brains of crustaceans with a characteristic feature is life-long neurogenesis (Sullivan and Beltz,2005). The brain (cerebral ganglion) plays a key role in research on replenishing neural progenitors(Hartenstein, 2014) and controlling sexual maturation(Tinikul et al., 2011) in crustaceans (Sullivan and Beltz, 2005; Lindsey and Tropepe, 2006; Stegner et al., 2015). Compared withDrosophila, the medulla,making up of about 40 thousand neurons, and representing in exceed of 70 kinds of cells, is the largest neuropil in the visual-processing center (optic lobe) (Li et al., 2013) using different labeling strategies. The functional important of different parts and neurons in redclaw crayfish remains little knowledge. Here, at least nine mainly different characteristic neurons are classified according to their morphological diversity and location inC.quadricarinatus. It has also mapped the distribution of astrocyte with GFAP labelling of the brain which agrees with what has previously been reported byProcambarusclarkia(de la Paz Rodríguez-Mu?oz and Escamilla-Chimal, 2015). Currently, it is still an urgent problem because of lacking more clear and definite markers to recognize different neurons.Hence, we should conduct research on the reactivity of cultured neurons to several types of antisera against the peptidergic hormones secreted from the brain and neurons’ secretory components to accurately classify the neurons and areas of brain (Hartenstein, 2006) to determine the function for each type of neuron in future. Maybe, the identification of such markers and their expression in brains of invertebrate animals promise to become a productive venue providing insight into neurons evolution.
As of right now, there is one of the key reasons why researchers failed to yield an obvious breakthough in analysis functions of gene and studying the neurophysiology due to lacking of crustaceans cell lines. Here, a method for primary cell culture with crayfish brain neurons was established inC.quadricarinatusin this report. With this culture system, it may be possible to determine the accurate location and function of the specific cell types in the brain ofC.quadricarinatusand understand the endocrine regulation at the cellular level. There is a possibility to achieve functional verification of genes by using primary cells in crayfish. Although our findings are required to achieve more efficient transfection of the primary cells, the method in primary cultured crayfish cells which were still unfacilitate in vitro for the expression of functional genes because of neurons quantitative limitation from CL9/11 and CL10. Compared with the whole brain’s neurons, maybe it’s just because there is existing activity of proliferative zones (MPZ and LPZ) with a high number of mitotically active cells (Sintoni et al.,2012). In the future, we need to improve yield rate of neurons from CL9/11 and CL10 under ensuring cell in good status to realize functional verification of genes.
5 CONCLUSION
The brain of crayfish is consist of three main parts.And at least 9 kinds of brain cells were identified through micro-examination. CL 9/11 and 10 cells could be used for primary culture about 7–9 d under the optimized L-15medium, but no cells were observed undergoing division. Maybe, it is difficult to establish a brain cell line for crayfish.This research provides a model that would benefit to investigate the cellular and synaptic mechanisms, sexual maturation,and molecular mechanisms of brain development because it shares similar structural characteristics with other crustaceans brain system (Stegner and Richter, 2011; Saetan et al., 2013; Ito et al., 2014;Stegner et al., 2015), such as layer structure and units.
6 ACKNOWLEDGEMENT
The authors are grateful to Prof. LIU Haipeng for cell culture training. We would like to give sincere thanks to Dr. GAO Yi and Dr. WANG Quanchao who helped to take some pictures in the manuscript.
7 AUTHOR CONTRIBUTION
H. D. and S. J. J. performed experiments and analyzed data. H. D, X. J. Z. F. H. L. and J. H. X.designed the experiments. H. D. wrote the manuscript,J. H. X. revised.
猜你喜歡
作物學(xué)報(2022年10期)2022-07-21 03:14:30
滇池(2022年4期)2022-03-24 01:43:42
小讀者(2021年8期)2021-11-24 05:59:50
小讀者·愛讀寫(2021年4期)2021-07-06 16:21:30
檢察風(fēng)云(2021年21期)2021-01-13 08:23:49
Communications in Theoretical Physics(2018年4期)2018-05-02 01:51:40
少年文藝·少年讀者文摘(2016年7期)2016-08-19 18:15:30
故事會(2014年21期)2014-05-14 15:24:23
 Journal of Oceanology and Limnology2018年6期
Journal of Oceanology and Limnology2018年6期
- Journal of Oceanology and Limnology的其它文章
- Effects of feeding time on complement component C7 expression in Pelteobagrus vachellii subject to bacterial challenge*
- Cryopreservation of strip spawned sperm using programmable freezing technique in the blue mussel Mytilus galloprovincialis*
- Pf- D mrt4, a potential factor in sexual development in the pearl oyster Pinctada f ucata*
- Specific genetic variation in two non-motile substrains of the model cyanobacterium Synechocystis sp. PCC 6803*
- Functional characterization of a Δ6 fatty acid desaturase gene and its 5′-upstream region cloned from the arachidonic acidrich microalga Myrmecia incisa Reisigl (Chlorophyta)*
- The expression characteristics of vitellogenin (VTG)in response to B(a)p exposure in polychaete Perinereis aibuhitensis*