
Genetic diversity assessment of a set of introduced mung bean accessions(Vigna radiata L.)
2018-04-12 03:33:59LixiWngPengBiXingxingYunHonglinChenSuhuWngXinChenXuzhenCheng
The Crop Journal 2018年2期
Lixi Wng,Peng Bi,Xingxing Yun,Honglin Chen,Suhu Wng,Xin Chen,Xuzhen Cheng,*
a Institute of Crop Sciences,Chinese Academy of Agricultural Sciences,Beijing 100081,China
b China National Seed Group Co.,Ltd.,Beijing 100031,China
c Institute of Vegetable Crops,Jiangsu Academy of Agricultural Sciences,Nanjing 210014,Jiangsu,China
1.Introduction
The mung bean(Vigna radiata L.),also known as green gram,is an important pulse crop providing vegetable protein for people throughout Asia[1].Mung bean is a traditional food in China and is widely grown in monoculture in dry and semi-dry regions,as well as being used as an intercrop throughout much of the country because of its drought tolerance and nitrogen-fixing soil fertilization[2].Owing to its short lifecycle,mung bean is also used as a post-disaster remedy when crops are destroyed by a natural disaster during their middle growth phase and it is too late to sow them again.However,the production of mung bean now faces novel challenges arising from climate change.The discovery of new genes and the development of varieties with multi-resistance or tolerance to diseases,pests,and extreme climates is the best strategy for sustainable production of this crop.
Assessments of phenotypic[3–5]or genotypic[6–8]diversity provide beneficial information for the better use of germplasm collections,which are the basic materialfor genetic improvement and breeding.Following a country-wide collection during the 1980′s,over 3000 accessions of mung bean germplasm were conserved in the national gene bank of China,and this collection is continually being added to[9–11].However,evaluations oflocal germplasm have shown that there are few elite genes conferring resistance to biotic and abiotic challenges,with the resultthatthe genetic backgrounds of modern varieties have become gradually narrowed[12].Previous reports have shown that alien or wild accessions can be of great value in local breeding[13,14].For instance,mung beans from the Asian Vegetable Research and Development Center(AVRDC)have contributed greatly to increasing the yields of local breeding cultivars in China compared to those at the end of the last century[12,15].Additionally,a bruchid-resistance gene has been successfully transferred to cultivated genotypes from wild mung beans[16,17].Thus,the best approach to enriching the local genetic pool is to introduce more germplasm from other countries or genes from wild relatives.
We recently obtained a set of mung bean accessions from the germplasm resources information center of the U.S.Department of Agriculture(USDA),a repository with a large mung bean collection.In the present study,we recorded the seed characters of these accessions,sowed them in Beijing to observe their growth,and evaluated their SSR variation.The main aims were 1)to determine whether the accessions can complete their lifecycle in Beijing and have potential use in cross-breeding without day length control,and 2)to assess the germplasm allelic richness by SSR variations to obtain information for further study.
2.Materials and methods
2.1.Plant materials
A total of 184 mung bean accessions,obtained from the germplasm resources information center of the USDA(http://www.ars-grin.gov/npgs/aboutgrin.html)in 2012,were used in the study.Of these accessions,178 were originally collected from 22 countries,two were from the Middle East,and four had no passport information(Table S1).After observation of their seed characters(color and testa),the accessions were planted in a greenhouse in early April 2013 in Beijing.Zhonglyu 5,a popular Chinese variety,was used as a control.Owing to the limited number of seeds available,only 10 seeds of each accession were planted,with row and plant spacing set at 50 cm and 12 cm,respectively.An equalweightoffresh leaves fromeach individual within each accession was collected for DNA extraction.
2.2.Phenotypic observation
In addition to seed characters,four other traits were investigated.Anthocyanidin coloration in the young stem was first observed and used to partially determine the homozygosity of each accession.Growth period,pod length,and seed weight were recorded as well,following Cheng et al.[18].These three characters provide information directly useful for the application of germplasm in local breeding programs.
2.3.SSR analysis
Genomic DNA was extracted using the CTAB method[19].Thirty-eight polymorphic SSR primers were used,with 24 from mung bean[20],12 fromadzukibean[21],and two fromcommon bean(obtained from http://www.ciat.cgiar.org/biotechnology/SSR_table.html).PCR analysis was performed in a 20-μL reaction solution containing 1× PCR buffer,100 μmol L?1of each dNTP,0.4 μmol L?1of each primer,20 ng genomic DNA,and 1 U of Taq DNA polymerase.Amplification was performed in an EDC-810 thermal cycler(Dongsheng Co.Beijing)with 35 cycles of 94°C for 30 s,47 °C for 30 s,and 72 °C for 30 s,followed by a final 5-min extension.The productwas separated by 8%SDS-polyacrylamide gel electrophoresis(PAGE),using 0.5×TBE as a buffer at 220 V.The running time of electrophoresis was adjusted according to the expected size of products,usually to 1.0–1.5 h.
2.4.Genetic diversity and STRUCTURE analysis
All SSR loci were scored as 1 if present and 0 if absent for each polymorphic fragment.The observed number of alleles(NA),the polymorphism information content(PIC value)and the expected heterozygosity were calculated using Popgene[22].
STRUCTURE 2.3.4[23,24]was applied based on multi-loci genotype data to assess the population structure of and the genetic relationships between the accessions.An admixture model and an independent allele frequency model were fitted to analyze data lacking prior population information.These models were run ten times for each number of populations(K)(varied from 1 to 10),and the maximum likelihood ratio was used to assign accessions to clusters[25].Genetic similarity coefficients(GSC)and genetic distances(GD)between accessions were calculated,and UPGMA(unweighted-pair-group method with arithmetic mean)was used for cluster analysis using NTSYS-pc2.10[26]based on genetic distances.
3.Results and analysis
3.1.Observations on the introduced germplasm
Among the 184 accessions,green seeds accounted for 78%and black seeds for 13%.Brown,yellow and dotted seed coats accounted for 3%,4%,and 3%,respectively.Seed testa with glossy surfaces accounted for 76%and those with dull surfaces for 22%.Three accessions were assumed to be mixed,because both glossy and dull seed testa were observed within one accession;of these,one was from India(PI 346316),one was from Pakistan(PI 268412),and the other had no passport information(PI363239).Thus,only seeds that had a prevalent seed testa were planted for the three accessions.
All of the accessions successfully emerged within a week after sowing.Eleven of them were observed to be genotype mixtures based on the anthocyanidin coloration of caulicles and cotyledons(Fig.1).Excluding the mixtures,most(79%)of the accessions showed anthocyanidin coloration.
Not all accessions completed their life cycles.Three accessions from India(PI 363489,PI 426128,and PI 363291)flowered but did not set seeds,while two(PI 427260 from Nepal and PI 363212 without passport information)did not flower.The growth periods of the 179 accessions varied from 82 to 121 days,with an average of 98.4 days.
Pod length varied from 4.4 cm to 15.8 cm with a mean of 7.3 cm.The original 100-seed weight varied from 0.82 g to 6.96 g with an average value of 3.42 g.There was a marked increase in the weights of newly harvested seeds,ranging from 1.64 to 7.30 with an average value of 3.87.Of the accessions,60%showed a 10%increase and 22%a decrease in seed weight.
Compared with Zhonglyu 5,a popular Chinese cultivar,some accessions showed extreme phenotypes for several traits little influenced by environment.Examples included a long-pod(15.8 cm)accession from India(PI 346314)and large-seeded(greater than 7.0 g per 100 seeds)accessions from India(PI363422,PI363316,and PI 363307),Korea(PI 362315),and China(PI 298915).
3.2.SSR genetic diversity
DNA samples were prepared from 184 accessions.A total of 159 polymorphic alleles at 38 SSR loci were observed,ranging from 3 to 6 alleles per locus with an average of 4.2.The number of alleles(NA)of SSR loci from adzuki and mung bean ranged from 3 to 6 with averages of 3.8 and 4.3,respectively.For SSR markers from common bean,3 and 5 alleles were detected for loci AG01 and PVBR201.Six polymorphic alleles were detected at 4 loci(CEDG008,Vr2-436,Vr3-429,and Vr3-598),5 at 9 loci,4 at 15 loci,and 3 at 10 loci(Table 1).The observed frequency of each allele for all 38 SSR loci by 184 accessions ranged from 0.5%to 78.2%.Eleven alleles had frequencies greater than 50.0%,while 28 alleles had frequencies lower than 5.0%,and unique alleles were observed at only 6 loci(Vr2-1063,Vr3-136,Vr3-429,Vr3-598,Vr3-671,and CEDC50).Accessions from India,Pakistan and Afghanistan displayed 99.0%of the total alleles.
The PIC values for 38 loci among the 184 accessions ranged from 0.365(CEDG144)to 0.753(Vr3-429),with an average of 0.650.The PIC values of the SSR derived from adzuki and mung bean varied from 0.365 to 0.713 and from 0.504 to 0.753,with averages of 0.608 and 0.665,respectively.AG01(0.735)and PVBR201(0.700)from common bean both showed relatively high values.
The expected heterozygosity ofthe 38 locivaried from0.357 to 0.751 with an average of 0.648.The expected heterozygosity for adzuki and mung bean SSRs varied from 0.357 to 0.711,and 0.504 to 0.751,with averages of 0.607 and 0.663,respectively.AG01(0.717)and PVBR201(0.694)from common bean both showed relatively high expected heterozygosity.
3.3.Genetic relationships and structure
The pairwise GSCs among the 184 accessions ranged from 0 to 0.96,with a mean value of 0.35.Five pairs showed GSCs of zero and 15 showed GSCs greater than 0.90.In total,more than 80%of the GSCs were lower than 0.5,but there were no clear trends in the distribution of GSCs between different countries(Fig.2).
STRUCTURE analysis based on the distribution of 159 alleles at 38 SSR loci showed that the 184 accessions could be divided into 6 groups(Fig.3),whereas three clusters were identified by NTSYS(Fig.S1).There was no distinct trend of accessions in different groups or clusters according to their origins.For example,accessions from India,Pakistan and Afghanistan were distributed among all six groups with no evident bias.
In total,group I was the smallest genetic group,consisting of 19 accessions from five Asian countries(India,Afghanistan,Pakistan,Iran,and Korea).Group II comprised 36 accessions:32 from Asia,2 from C?te d'Ivoire(PI 286298,PI 286306),one from Australia(PI 426046)and the other of unknown origin(PI 363272).The origins of accessions in group III(35 accessions),group IV(35 accessions)and group V(25 accessions)were widely distributed,and the four accessions without passport information were separately placed into groups II to V.Group VI comprised 34 accessions and most of them were also from Asia,except one from Guatemala(PI 227754),which is in Central America.

Fig.1–Anthocyanidin coloration in the caulicle and cotyledon of mung bean.A,coloration in the cotyledon;B,coloration in the caulicle.

Table 1–Observed number of alleles (NA)and polymorphism information content determined in 184 accessions of mung bean using SSR markers.
4.Discussion
4.1.Management of germplasm collections
Mixed accessions complicate the management and use of germplasm.There have been several studies of mung bean genetic diversity using phenotypes[3,27,28]and genotypes[29–33],but few have mentioned the phenomenon of accession mixtures in genetic resources that we observed based on both anthocyanidin coloration and seed coat luster.Anthocyanidin coloration,controlled by a dominant single gene[34],is a simple and obvious character used to determine at the seedling stage whether an accession is a mixture.Anthocyanidin coloration has been listed as one of the basic characters of mung bean cultivars or accessions[12]and is often used to purify cultivars at the seedling stage.Reports of the inheritance of seed coat luster differ,suggesting a dominant single gene[4,35]or two genes[36,37].More complex genetic mechanisms have also been recently proposed[38].However,seed coat luster is stable under different environments and is often used as a cultivar character.
4.2.The performance of introduced mung bean germplasm in China
The normal growth of alien germplasm is a basic requirement for its use in local breeding programs.In the present study,all of the introduced mung bean accessions completed their lifecycles,indicating that we can apply them into breeding without day length control.For those that could not set seeds or even could not flower,extra management should be applied before using.
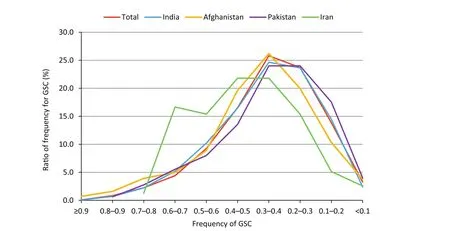
Fig.2–Distribution of frequencies of GSC among 184 introduced mung bean accessions based on 38 SSR markers.
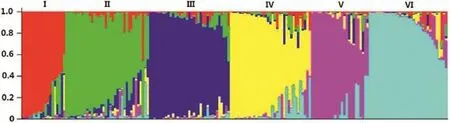
Fig.3–Genetic structure of 184 introduced mung bean accessions inferred from 159 SSR alleles.
The long growth period of 82–121 days observed in the present study is attributable to the early April sowing date,given that Zhonglyu 5,a cultivar with a growth period of no more than 70 days when sown around June in Beijing,also had a relatively long growth period(85 days)in this study.
We identified accessions with other obvious beneficial characters,such as an erect habit,dwarfed plants,and increased pods per plant.These characters should be further validated,given that the numbers of seeds of the introduced accessions were limited and no field replications were performed.However,observations oftraits thatwere moderately stable under different environments were reliable.For example,pod length and large seed are both associated with high yield and have been used as breeding objectives for mung beans.One accession had a long pod(15.8 cm)and three had a large seed(at least 7 g per 100 seeds),phenotypes more extreme than those of the local collection[9–11].These accessions can be used in local breeding programs by crossing with local cultivars and might broaden the genetic backgrounds of cultivars.
4.3.SSR genetic diversity and backgrounds
In contrast to previous studies on mung bean diversity analysis with SSR markers,we observed a higher diversity level,considering the distribution of the germplasm studied(Table 1).In total,the NA ranged from 2 to 6 in most of the diversity analyses of the mung bean[6,7,29,30,32,39,40]as we list in Table 2,and only an average of 6.1 alleles were detected within a core collection of 1481 mung bean accessions[7],indicating low genetic variation in the mung bean compared to the soybean genome[41].We detected an average of 4.2 alleles per locus using 184 accessions,a value similar to that detected in 692 accessions from 27 countries[30]and higher than that detected in 157 accessions using EST-SSR markers[29].We also detected a higher average PIC value compared to those in previous reports(Table 2),indicating a greater richness in the distribution of alleles.The GSCs in the present study were in agreement with the calculated diversity values.Most of the GSCs were lower than 0.50 and there were some accession pairs with GSCs of zero,suggesting that they had completely different genetic backgrounds.
The population structures inferred by STRUCTURE and NTSYS methods were not in complete agreement.However,some clear trends appear(Fig.S1).For example,most of the accessions in group I and III were divided into cluster III,but in sub cluster II and I,respectively.Accessions in groups IV and V,were found in respectively sub cluster I and II of cluster II.Neither method grouped the accessions according to theirorigins,nor the accessions from other continents distributed regularly into the Asian groups.Even accessions from the same country formed different groups.The disagreement between these results and those of previous studies in mung bean[33]and related species[42],might be attributed to several factors.First,sample size affects the accurate assessment of genetic background,and the distribution of accessions from different countries and regions in the present study was uneven.Second,after a long period of cultivation,accessions may adapt to local conditions,whereas those maintained in collections are preserved in the gene bank almost constantly,except for reproduction,so that they are rarely exposed to variations in the local environment.Third,because seed is frequently transported between countries or agencies,some original passport information of accessions might be misregistered,affecting our estimation of their relationships.This circumstance also suggests that introduced accessions should always retain their original codes in both collection and introduction to avoid extra labor in germplasm management.

Table 2–Comparison of genetic diversity detected within different mung bean germplasm by SSR markers.
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2017.08.004.
This work was supported by the China Agriculture Research System(CARS-09),the Agricultural Science and Technology Innovation Program(ASTIP)of CAAS and the National Key Technology R&D Program of China(2013BAD01B03-18).
[1]N.Tomooka,D.Vaughan,H.Moss,N.Maxted,The Asian Vigna:Genus Vigna Subgenus Ceratotropis Genetic Resources,Kluwer Academic Publishers,London,UK,2003.
[2]Z.J.Zheng,Food Legumes in China,Chinese Agriculture Press,Beijing,China,1995(in Chinese).
[3]I.S.Bisht,R.K.Mahajan,D.P.Patel,The use of characterization data to establish the Indian mung bean core collection and assessment of genetic diversity,Genet.Resour.Crop Evol.45(1998)127–133.
[4]L.X.Wang,X.Z.Cheng,S.H.Wang,J.B.Li,J.X.Li,Genetic variability of agronomic traits in mung bean applied core collection of China,J.Plant Genet.Resour.10(2009)589–593(in Chinese with English abstract).
[5]F.Abna,F.Golam,S.Bhassu,Estimation of genetic diversity of mung bean(Vigna radiata L.Wilczek)in Malaysian tropical environment,Afr.J.Microbiol.Res.6(2012)1770–1775.
[6]P.Somta,W.Musch,B.Kongsamai,S.Chanprame,S.Nakasathien,T.Toojinda,W.Sorajjapinun,W.Seehalak,S.Tragoonrung,P.Srinives,New microsatellite markers isolated from mung bean(Vigna radiata(L.)Wilczek),Mol.Ecol.Resour.8(2008)1155–1157.
[7]R.Schafleitner,R.M.Nair,A.Rathore,Y.W.Wang,C.Y.Lin,S.H.Chu,P.Y.Lin,J.C.Chang,A.W.Ebert,The AVRDC-the world vegetable center mung bean(Vigna radiata)core and mini core collections,BMC Genomics 16(2015)344.
[8]P.Lestari,S.K.Kim,Reflinur,Y.J.Kang,N.Dewi,S.Lee,Genetic diversity of mung bean(Vigna radiata L.)germplasm in Indonesia,Plant Genet.Resour.12(2014)S91–S94.
[9]Z.J.Zheng,Catalogues of Chinese Food Legumes Germplasm and Resources:Continuation I,China Agriculture Press,Beijing,China,1987(in Chinese).
[10]Z.J.Zheng,Catalogues of Chinese Food Legumes Germplasm and Resources:Continuation II,China Agriculture Press,Beijing,China,1990(in Chinese).
[11]J.P.Hu,X.Z.Cheng,P.Z.Wang,Catalogues of Chinese Food Legumes Germplasm and Resources:Continuation III,China Agriculture Press,Beijing,China,1996(in Chinese).
[12]X.Z.Cheng,S.M.Wang,The History of Chinese Food Legume Varieties,China Agricultural Science and Technology Press,Beijing,China,2009(in Chinese).
[13]N.Tomooka,C.Lairungreang,P.Nakeeraks,Y.Egawa,C.Thavarasook,Development of bruchid-resistant mung bean line using wild mung bean germplasm in Thailand,Plant Breed.109(1992)60–66.
[14]Y.H.Lee,J.K.Moon,K.Y.Park,J.H.Ku,H.T.Yun,W.K.Chung,S.D.Kim,H.S.Kim,D.H.Kim,M.N.Chung,A new mung bean cultivar with bruchid resistance,‘Jangannogdu',Korean J.Breed.32(2000)296–297.
[15]X.L.Tong,S.Xu,G.H.Li,H.J.Zhang,Q.N.Wang,Studies on sowing test and introduction of mung bean from AVRDC,Rain Fed Crops 22(2002)355–356(in Chinese).
[16]H.J.Zhu,X.Y.Zhao,H.B.Yan,W.Gao,Y.W.Zhang,Breeding of bruchid resistant mung bean variety Jinlyudou 7,J.Shanxi Agric.Sci.40(2012)606–607(in Chinese with English abstract).
[17]H.T.Chen,X.X.Yuan,H.M.Zhang,X.Q.Liu,X.Y.Cui,X.Chen,H.P.Gu,Breeding and cultivation techniques of the new mung bean variety Sulyu no.5 with bruchid resistance,Crop Res.29(2015)428–430(in Chinese).
[18]X.Z.Cheng,S.H.Wang,L.X.Wang,Descriptors and Data Standard for Mungbean(Vigna radiata(L.)Wilczek),China Agricultural Science and Technology Press,Beijing,China,2006(in Chinese).
[19]J.J.Doyle,J.L.Doyle,A rapid DNA isolation procedure for small quantities of fresh leaf tissue,Phytochem.Bull.19(1987)11–15.
[20]L.X.Wang,M.El Baidouri,B.Abernathy,H.L.Chen,S.H.Wang,S.H.Lee,S.A.Jackson,X.Z.Cheng,Distribution and analysis of SSR in mung bean(Vigna radiata L.)genome based on an SSR-enriched library,Mol.Breed.35(2015)25.
[21]O.K.Han,A.Kaga,T.Isemura,X.W.Wang,N.Tomooka,D.A.Vaughan,A genetic linkage map for azuki bean[Vigna angularis(Willd.)Ohwi&Ohashi],Theor.Appl.Genet.111(2005)1278–1287.
[22]F.C.Yeh,T.J.B.Boyle,Population genetic analysis of codominant and dominant markers and quantitative traits,Belg.J.Bot.129(1997)157.
[23]J.K.Pritchard,M.Stephens,P.Donnelly,Inference of population structure using multilocus genotype data,Genetics 155(2000)945–959.
[24]N.A.Rosenberg,J.K.Prichard,J.L.Weber,H.M.Cann,K.K.Kidd,L.A.Zhivotovsky,M.W.Feldman,Genetic structure of human populations,Science 298(2002)2381–2385.
[25]G.Evanno,S.Regnaut,J.Goudet,Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study,Mol.Ecol.14(2005)2611–2620.
[26]F.J.Rohlf,NTSYS-pc Numerical Taxonomy and Multivariate Analysis System,State University of New York,New York,USA,1992.
[27]L.X.Wang,X.Z.Cheng,S.H.Wang,X.Zhu,Z.X.Liu,Adaptability and phenotypic variation of agronomic traits in mung bean core collection under different environments in China,Acta Agron.Sin.40(2014)739–744(in Chinese with English abstract).
[28]L.Qiao,H.L.Chen,L.X.Wang,S.H.Wang,X.Z.Cheng,Y.W.Zhang,Genetic diversity of foreign mung bean germplasm resources by agronomic characters,J.Plant Genet.Resour.16(2015)986–993(in Chinese with English abstract).
[29]H.L.Chen,L.Qiao,L.X.Wang,S.H.Wang,M.W.Blair,X.Z.Cheng,Assessment of genetic diversity and population structure of mung bean(Vigna radiata)germplasm using EST-based and genomic SSR markers,Gene 566(2015)175–183.
[30]J.G.Gwag,A.Dixit,Y.J.Park,K.H.Ma,S.J.Kwon,G.T.Cho,G.A.Lee,S.Y.Lee,H.K.Kang,S.H.Lee,Assessment of genetic diversity and population structure in mung bean,Genes Genomics 32(2010)299–308.
[31]G.R.Lavanya,J.Srivastava,S.Ramade,Molecular assessment of genetic diversity in mung bean germplasm,J.Genet.87(2008)65–74.
[32]D.Shrivastava,P.Verma,S.Bhatia,Expanding the repertoire of microsatellite markers for polymorphism studies in Indian accessions of mung bean(Vigna radiata L.Wilczek),Mol.Biol.Rep.41(2014)5669–5680.
[33]Y.Liu,X.Z.Cheng,L.X.Wang,S.H.Wang,P.Bai,C.S.Wu,Genetic diversity research of mung bean germplasm resources by SSR markers in China,Sci.Agric.Sin.46(2013)4197–4209(in Chinese with English abstract).
[34]L.X.Wang,X.Z.Cheng,S.H.Wang,Y.Liu,Inheritance of several traits in mung bean(Vigna radiata),Acta Agron.Sin.39(2013)1172–1178(in Chinese with English abstract).
[35]H.A.Rheenen,The inheritance of some characters in green gram,Genetika 36(1966)412–419.
[36]P.H.Manguiat,R.M.Lantican,A.L.Carpena,I.G.Catedral,Inheritance of seed color,size,and luster in mung bean(Vigna radiata L.Wilczek),Philipp,J.Crop.Sci.3(1978)238–241.
[37]S.K.Bhadra,M.L.Akhter,A.Quadhem,Genetics of seed lustre and joint inheritance of seed-coat color and seed lustre in mung bean,Bangladesh J.Bot.20(1991)61–64.
[38]C.J.Lambrides,I.D.Godwin,R.J.Lawn,B.C.Imrie,Segregation distortion for seed testa color in mung bean(Vigna radiata L.Wilczek),J.Hered.95(2004)532–535.
[39]S.Tangphatsornruang,P.Somta,P.Uthaipaisanwong,J.Chanprasert,D.Sangsrakru,W.Seehalak,W.Sommanas,S.Tragoonrung,P.Srinives,Characterization of microsatellites and gene contents from genome shotgun sequences of mung bean(Vigna radiata(L.)Wilczek),BMC Plant Biol.9(2009)137.
[40]P.Somta,W.Sommanas,P.Srinives,Molecular diversity assessment of AVRDC-the world vegetable center eliteparental mung bean,Breed.Sci.59(2009)149–157.
[41]L.X.Wang,R.X.Guan,Z.X.Liu,R.Z.Chang,L.J.Qiu,Genetic diversity of Chinese cultivated soybean revealed by SSR markers,Crop Sci.46(2006)1032–1038.
[42]A.Asfaw,M.W.Blair,C.Almekinders,Genetic diversity and population structure of common bean(Phaseolus vulgaris L.)landraces from the East African highlands,Theor.Appl.Genet.120(2009)1–12.

- The Crop Journal的其它文章
- Near-infrared reflectance spectroscopy reveals wide variation in major components of sesame seeds from Africa and Asia
- Alternate phenotype–genotype selection for developing superior high-yielding irrigated rice lines
- F unction of the auxin-responsive gene TaSAUR75 under salt and drought stress
- Development and validation of simple sequence repeat markers from Arachis hypogaea transcript sequences
- Soybean hairy roots produced in vitro by Agrobacterium rhizogenes-mediated transformation
- Elevated temperature intensity, timing, and duration of exposure affect soybean internode elongation, mainstem node number, and pod number per plant