
Genetic variation and selection of introduced provenances of Siberian Pine (Pinus sibirica) in frigid regions of the Greater Xing’an Range, Northeast China
2014-04-20 06:58:00XiyangZhaoChengWangShuchunLiWeiHouShiqingZhangGuijieHanDengPanPingWangYufeiChengGuifengLiu
Journal of Forestry Research 2014年3期
Xiyang Zhao · Cheng Wang · Shuchun Li · Wei Hou · Shiqing Zhang Guijie Han · Deng Pan · Ping Wang · Yufei Cheng · Guifeng Liu
Introduction
Siberian Pine (Pinus sibirica) is an extremely hardy, long-lived and cold-resistant species, and is widely distributed in pristine forests throughout northern Russia. For centuries, P. sibirica has been used by native Siberians as an adaptable source of valuable timber, natural remedies, and edible pine nuts. The bark, needles, resin, nutshells and pine nut kernels have been widely used for their unique disease healing properties (Krylov et al. 1983). Recent studies showed that nuts of P. sibirica had outstanding nutritional, health- supporting properties and nut oil (Alantsev 1981; Isaev 1985; Krylov et al. 1983).
Due to the extreme environment in northeast China, there are few plant species in most alpine forests and it is therefore difficult to meet the demands of environment protection and national economy. Especially in the frigid zone of the Greater Xing’an Range, the scarcity of useful tree species is the most serious problem for the forest industry. Since the environment of the Greater Xing’an Range is similar to the natural range of P. sibirica, four provenances of P. sibirica were introduced from Mongolia and Russia in 1993 and young trees were planted after the seedlings were cultivated for three years. Introduction of P. sibirica was carried out for 20 years to 2013. This study focused on the genetic variation in survival rates and growth traits of P. sibirica 18 years after planting, and the status of tree height from year 11 to year 18. Results of this study provide a new theoretical basis for future work.
Materials and methods
Experiment site
The introduction experiment was carried out at the Bizhou Forestry Centre (Forestry Bureau of Xinlin), which is located on the north slope of Yin Le Shout Mountain in the Greater Xing’an Range in northeast China (124o40' E, 51°43' N ) and the elevation was 1000 m above mean sea level. The climate of this area was classified as cold temperate monsoon, and it is cold and dry in winter, with little rainfall in late spring and early summer. Rainfall was concentrated in July and August, and average annual rainfall was 473 mm. Average annual temperature was -4°C and the recorded lowest temperature in winter was -52°C. The soils at the experiment site were lean, thin, and short of nutrients.
Materials and experimental design
Based on the climate and geographical characteristics of the experimental site, seeds of four P. sibirica provenances were obtained from four sites: Tomsk, Novosibirsk, Altai Mountain in Russia, and North Hanggay in Mongolia (Fig. 1). The climatic factors of these locations are listed in Table 1. The seeds were germinated in 1993 in greenhouse and the seedlings were planted at the experimental site after two years. The experiment utilized a randomized complete block design (Marron et al. 2006), with six blocks containing 25 seedlings of the same provenance in each square plot (5 m × 5 m). Tree seedlings were planted in rows with a 2.0 m × 1.5 m spacing for each block.
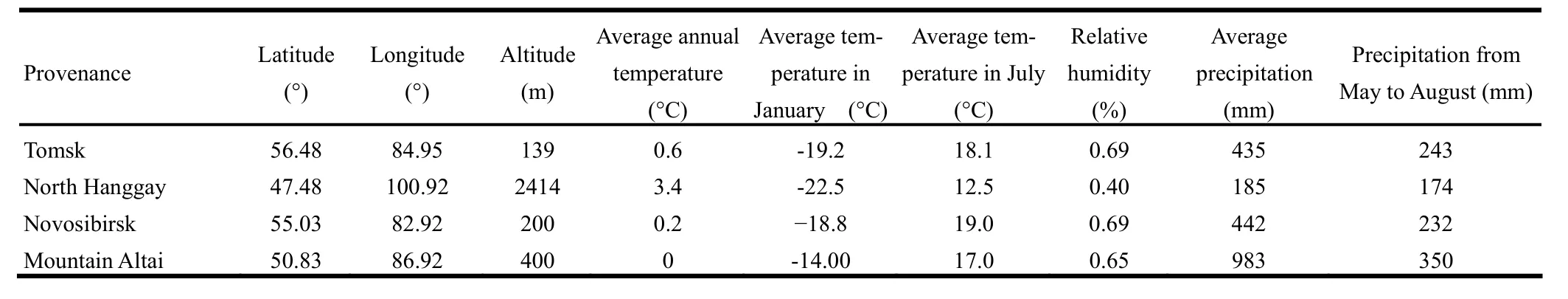
Table 1: Geographical and climatic factors for the four P. sibirica provenances
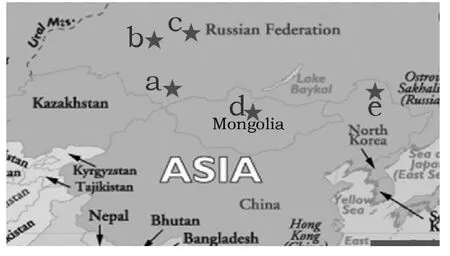
Fig. 1: Collection sites of the four P. sibirica provenances and location of the experiment which were denoted by red stars, site a: Mountain Altai; b: Novosibirsk; c: Tomsk; d: North Hanggay; e: Bizhou (the experimental site in northeastern China).
Data collection
From growth years 11 to 18, the height (H) of each tree was measured. At the year 18, basal diameter (BD) and crown length (CL) were also measured for all normal-growth trees (data for dead or broken trees were omitted). Survival rates were also measured for each of the four provenances.
Statistical analyses
Statistical analyses were carried out using SPSS version 13.0. The significance of fixed effects was tested using the F test. Variation in tree height was analyzed by the following model (Hansen et al. 1996):
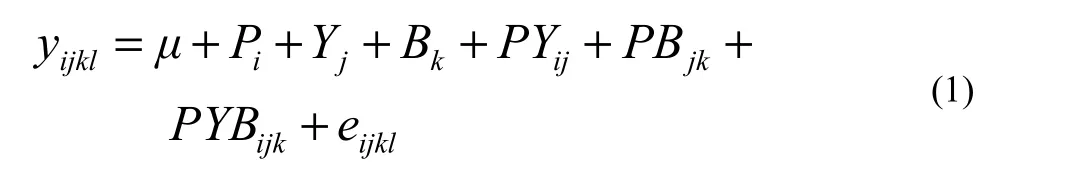
where yijklis the observation on the lth ramet from the ith provenance of the jth year in the kth block; μ is the overall mean; Piis the effect of the ith provenance (i = 1,……4); Yjis the effect due to the jth year (j = 11,……18); Bkis the effect of the kth block (k = 1,……6); PYijis the interaction between the ith provenance and jth year, PBjkis the interaction between the jth provenance and kth block, the PYBijkis the interaction among the ith provenance, the jth year and the kth block, and eijklis the random error.
Variation among provenance of the samples was analyzed using analysis of variance (ANOVA) within sites (Hansen et al. 1996):
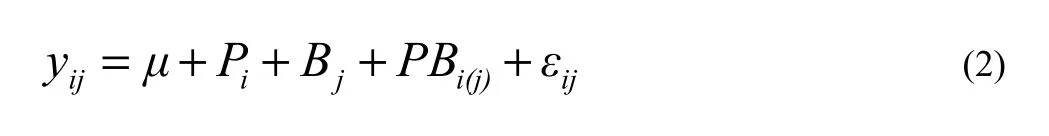
where yijis the performance of the ramet of the ith provenance within the jth block, μ is the overall mean; Piis the effect of the ith provenance (i =1,……4); Bjis the effect of the jth block (i = 1,……6); PBi(j)is the random effect of the ith provenance within the jth block and εijis the random error.
The coefficient of phenotypic variation (PCV) for provenance was calculated using the following formula (Hai et al. 2008):

Individual repeatability (H2; Hansen et al. 1996) was calculated as:
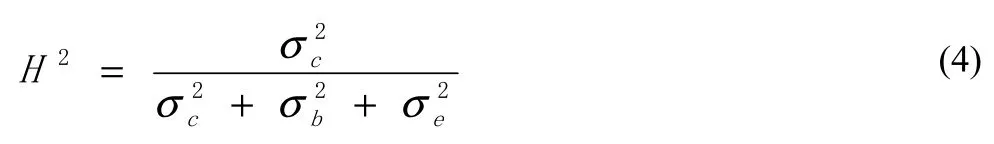
The phenotype correlation rA(xy) of traits x and y was calculated as:
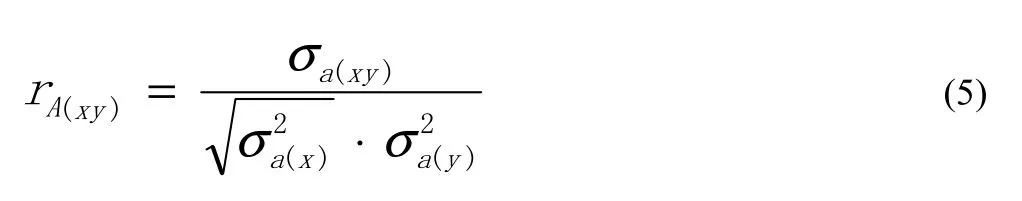
The colligation assessment was calculated as:
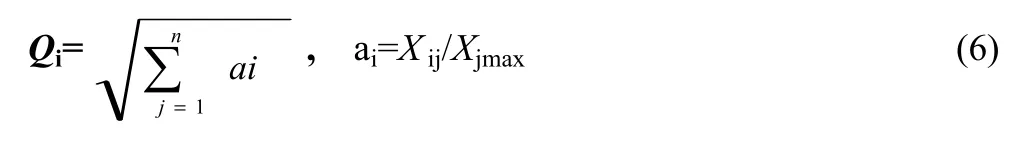
where Qiis the value of colligation assessment, Xijis an average value of one trait, Xjmaxis the maximum of the trait, and n is the trait number (Zhao et al. 2012).
Results
Variation in H from year 11 to year 18 after planting
ANOVA analysis for H across growth years is presented in Table 2. Variations by provenance and growth year were both highly significant (p <0.01). The interaction between provenance and growth year was significant (p <0.05).
Genetic and variation parameters for different provenances
ANOVA analyses for growth traits of P. sibirica are shown in Table 3. All traits, including survival rate, varied significantly between the four provenances (p < 0.01). The average survival rate at year 18 was 83.59%. Average tree height of all trees at year 11 was 114.90 cm, and up to year 18, average tree height increased to 349.37 cm. At year 18, average BD was 4.11 cm and average CL was 60.11 cm. H2and PCV of each trait are also shown in Table 3. H2values for tree height were high, varying from 0.967 to 0.983, and increased as trees grew. Traits BD and CL also had high H2values at year 18, with values of 0.974 and 0.968 respectively. As trees grew older, PCV for H among different provenances also increased. PCV for BD and CL at year 18 were 37.60% and 31.44% respectively. High heritability and PCV suggested that broad variation existed between provenances and that the variation was strongly controlled by genetic factors. These results were suitable for selecting the most suitable provenances.
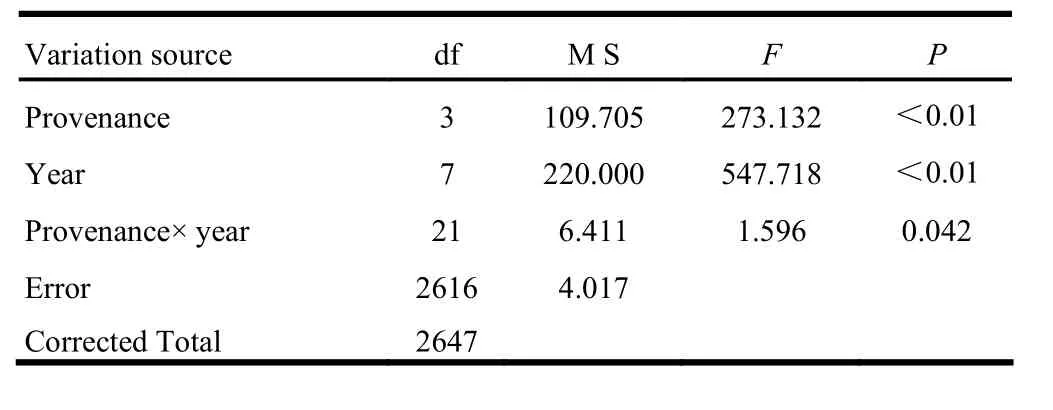
Table 2: ANOVA analysis of tree height for the four P. sibirica provenances from the 11 to 18 growth years
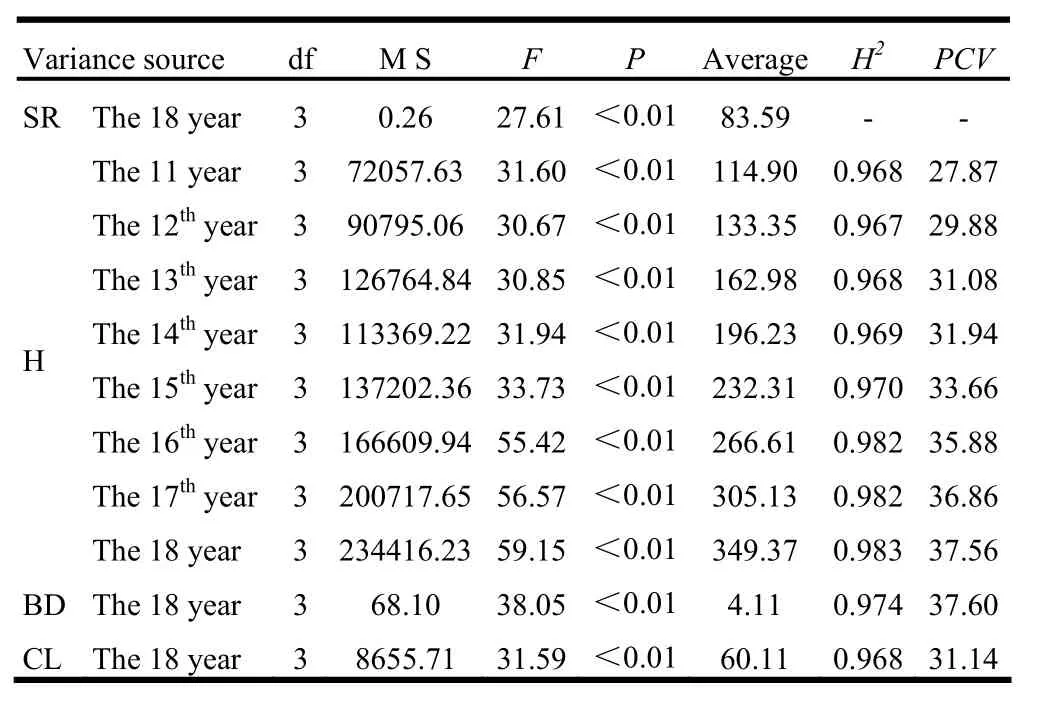
Table 3: ANOVA analysis and genetic parameters for different traits of the four P. sibirica provenances
Survival rate, H, BD and CL of the four provenances at growth year 18
Survival rates of each P. sibirica provenance are shown in Fig. 2. The survival rate of the Novosibirsk provenance was 95.33%, which was the highest of the four provenances. Following this, the Altai Mountain provenance (93.67%) and the Tomsk provenance (92.67%) also indicated high survival rates. North Hanggay provenance had a lower survival rate of 52.67%.
Growth traits (H, BD and CL) of the four P. sibirica provenances at growth year 19 are displayed in Figs. 2b, c, and d, respectively. The Tomsk provenance had the highest average H (384.11 cm) and BD (4.65 cm), while the Novosibirsk provenance showed the same BD as that of Tomsk provenance but the largest CL (65.77 cm) of the four introduced provenances. Compared with the other three provenances, the North Hanggay provenance had a considerably lower production.
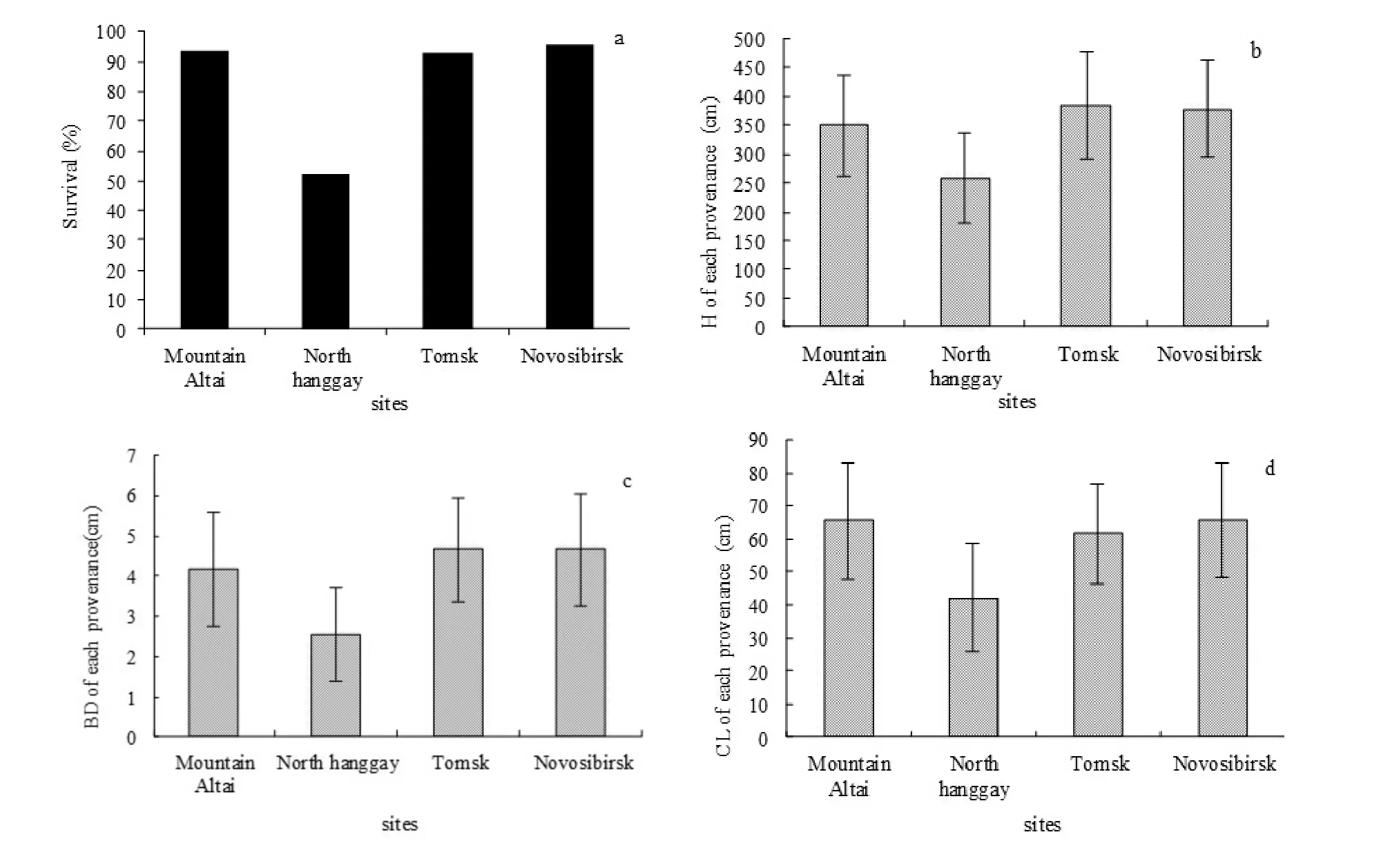
Fig. 2: Survival rate (a), H (b), BD (c) and CL (d) for the four P. sibirica provenances at the 18 year following introduction to China. Survival rate, height, basic diameter, and tree canopy length showed that the Novosibirsk introduction was superior to the others, and that the North Hanggay provenance showed the poorest performance.
Average H and PCV of provenances from growth year 11 to 18
The growth curves for trees from the year 11 to 18 are shown in Fig. 3. At the year 11, the Tomsk provenance showed the highest H (135.79 cm) of the four provenances. The North Hanggay provenance showed the lowest H. At year 18, the Tomsk provenance maintained the highest growth rate, and showed the highest H (384.11 cm). The North Hanggay provenance showed the lowest growth rate and at year 18, the average H was 258.29 cm.
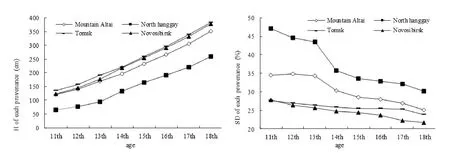
Fig. 3: H and PCV for each P. sibirica provenance from the 11 to 18 growth year. Tree heights were increased but PCV of height gradually decreased.
PCVs for height from year 11 to 18 are shown in Fig. 3. At the year 11, PCV for the North Hanggay provenance (47.12%) was higher than for other provenances. PCV of the Altai Mountain provenance was somewhat lower than that of the North Hanggay provenance. The Tomsk and Novosibirsk provenances showed the lowest PCV values. PCV decreased with increasing age for all provenances. At growth year 18, PCVs for the North Hanggay and Novosibirsk provenances decreased significantly to 30.11% and 21.6% respectively.
Correlation coefficients among all traits
Correlation coefficients among growth traits are shown in Table 4. All correlation coefficients were positive, significant and higher than 0.900. From year 11 to 18, the correlation coefficients of H varied from 0.967 to 0.999. The correlation coefficients of H increased with tree growth. At year 18, BD and CL were positively correlated with H, and the coefficient between H and BD reached 0.986; coefficients between CL and H, and CL and BD were 0.965 and 0.964 respectively.
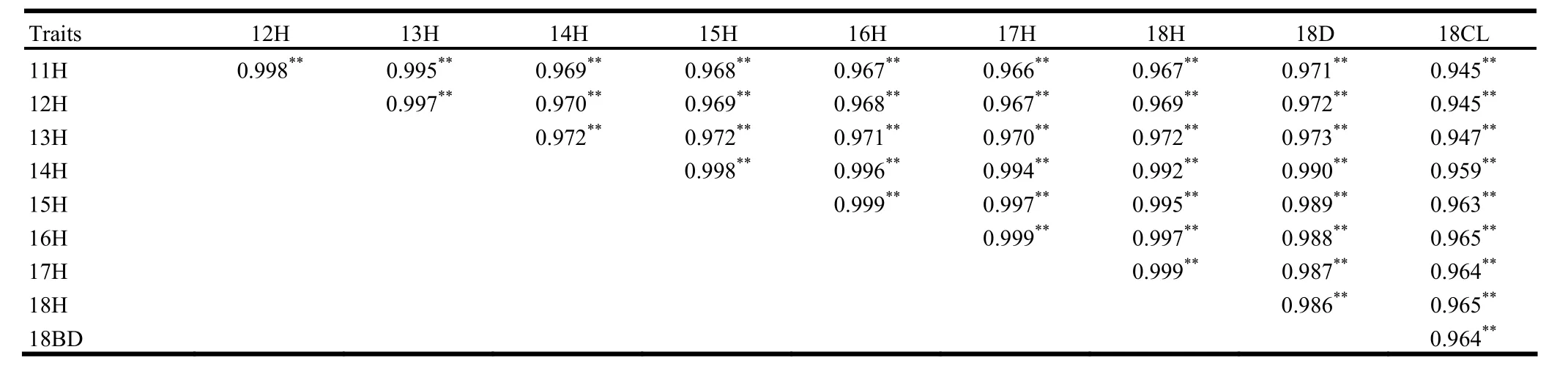
Table 4: Correlation coefficients for growth traits of the four P. sibirica provenances.
Correlation coefficients between growth traits and environmental factors
The original habitats of the four P. sibirica provenances were widely separated and differed greatly in geography and climate. Primary geographical differences included latitude, longitude and elevation. Main climatic factors were average annual temperature, maximum and minimum temperatures, relative humidity, and average precipitation. The correlation coefficients between environmental and growth parameters are shown in Table 5. All coefficients were significant (p <0.01) which indicated the original environmental factors significantly influenced plant growth. Longitude, elevation, and average annual temperature were negatively correlated but latitude and other environment factors were positively correlated with growth traits. The correlation analysis of growth traits at year 18 with environments of plantation sites are shown in Table 6. Similar to the source regions, all correlations were significant (p <0.01). H, BD and CL were positively correlated with latitude, temperature and precipitation, but negatively correlated with longitude and elevation. Most correlation coefficients for the plantation sites were higher in absolute value than the source sites, indicating that the influence of the source environment declined as trees grew and trees gradually adapted to the new environment.
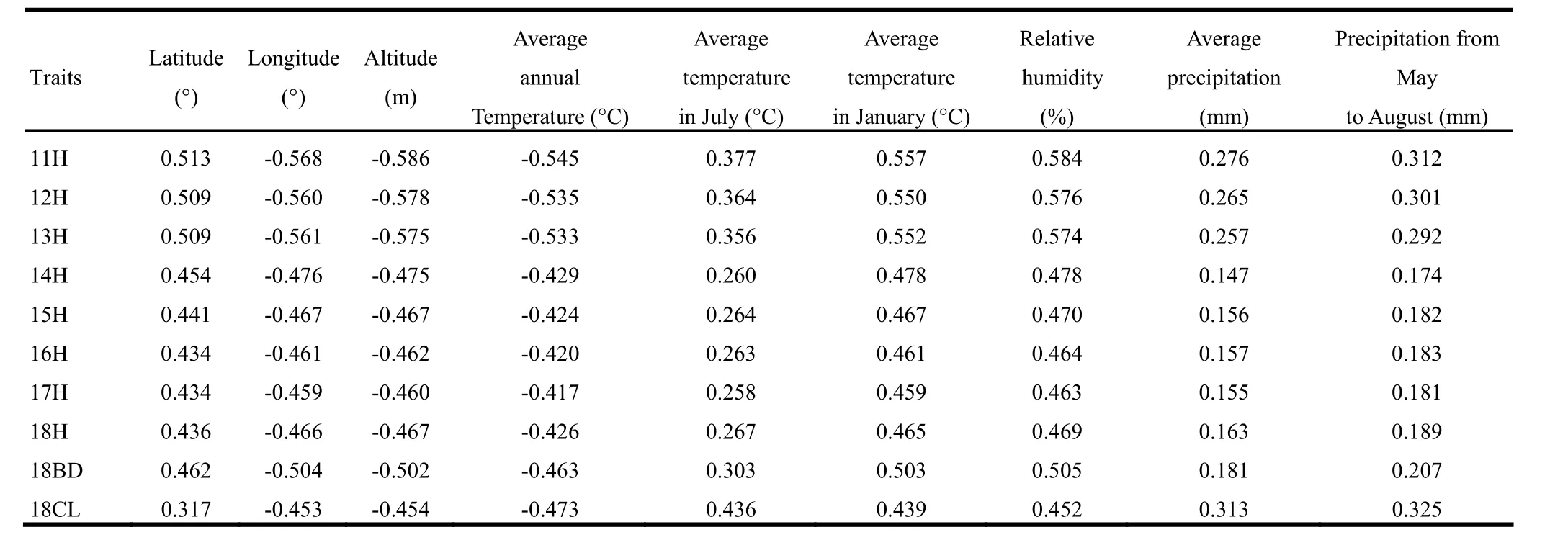
Table 5: Correlation coefficients between growth traits and original environmental factors
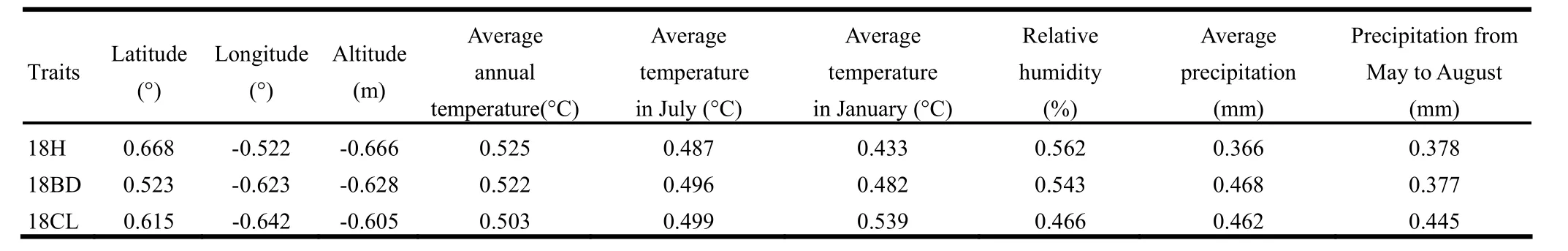
Table 6: Correlation coefficients between growth traits and environmental factors in the test site at the 18 growth year
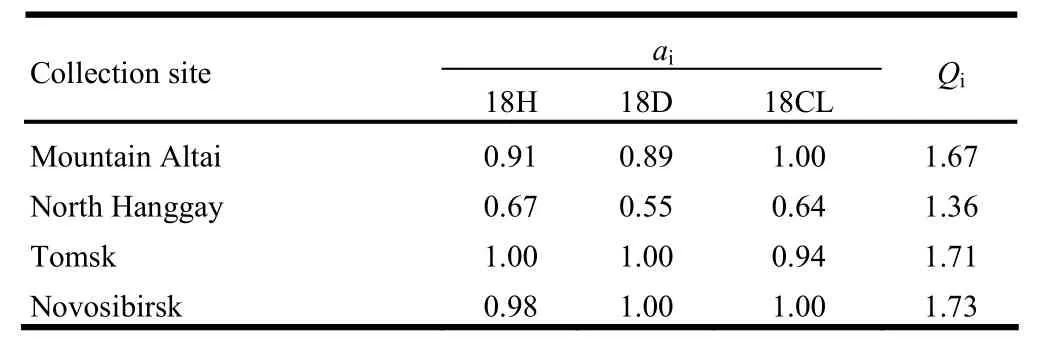
Table 7: Qi values for the four P. sibirica provenances
Colligation estimates of introduction results
Methods for tree selection rely on the breeding objective of tree genetics and breeding. P. sibirica is an important species with multiple uses for food, medicinal resources, timber and also ecological demand. Therefore multiple traits should be considered simultaneously in provenance selection. In this experiment, H, BD and CL at growth year 18 were colligated to estimate a Qivalue for each provenance (Table 7). The Qivalue of the Novosibirsk provenance was the highest of the four provenances (Qi=1.73), followed by the Tomsk provenance (Qi=1.71) and the Altai Mountain provenance (Qi=1.67). The North Hanggay provenance had the lowest Qivalue of 1.36.
Discussion
Humans have long moved plants into new ranges or habitats. The primary reasons for the intentional transport of plants have been for agriculture (food or forage) and for medicinal uses. Plant introductions release alien species into environments in which they were not indigenous, and most staple crops (e.g. maize, potatoes, rice, wheat) are cultivated far from their native ranges. Introductions have historically been followed by selection for varieties that were adapted to survival and yield under local conditions. In addition to edible crops, many species of woody plants have been transported and established in new ranges either as ornamentals or to provide a commodity such as lumber or rubber. Of great concern to ecologists is the potential for an introduced exotic species to escape cultivation and become invasive, with the potential to cause profound changes to the environment (e.g. loss of biodiversity, hybridization with native species, introduction of exotic pests or pathogens, changes to local water use patterns) as well as economic damage (Engelmark et al. 2001; Peterken 2001). For trees, which may have very long lifespans, deleterious issues directly related to introduction may not become apparent for many decades (Peterken 2001). Fortunately, the rate of spread of naturalized exotic trees is typically only meters/year (Mack and Lonsdale 2001). Practical experience in Sweden showed that successful large-scale introduction of lodgepole pine (Pinus contorta var. latifolia) required extensive planning, management and monitoring to reduce exotic species’effects on biodiversity and the environment (Engelmark et al. 2001).
In this study, we examined growth traits and survival rates of four distinct provenances of Siberian Pine (P. sibirica) planted in the Greater Xing’an Range in northeast China. This species was well suited for introduction studies in China because it has an extensive natural range (>36 million hectares in northeast Russia, the Ural Mountains and Siberia, as well as populations in northern Mongolia) and could be resistant to white pine blister rust caused by the fungus Cronartium ribicola. Long-term studies in Russia showed that there existed wide variation in P. sibirica and the seeds originating from lower elevations in southern Siberian mountains grew faster than those from northern provenances (Iroshnikov and Politov 2004). Our results showed that the Novosibirsk, Tomsk, and Altai Mountain provenances showed high survival rates and high biomass production, and were therefore suitable for introduction into the Greater Xing’an Range in northeast China.
Trait-based field studies have shown that considerable intraspecific genetic variation exists between provenances in P. sibirica (Iroshnikov and Politov 2004), as well as in P. sylvestris, P. radiata and P. pinea (Danusevi?ius 2001; Wu et al. 2007; Carrasquinho and Gon?alves 2013). For Siberian pine, studies using allelic variation in isozymes have also shown a high level of genetic variation for the species (summarized in Politov and Krutovsky 2004). In this experiment, there were significant differences not only between provenance and age, but also in the interaction of age × provenance in tree height. Within all growth years, variations for height were significant, as were differences in BD and CL at year 18. These findings indicate that there was considerable variation in growth traits among the four P. sibirica provenances, and these results were favorable for genetic selection. Analysis of variation in survival rate, height, basal diameter, and tree canopy length showed that the Novosibirsk provenance was superior to the others, and the North Hanggay provenance showed the poorest performance.
Natural genetic variation in undomesticated populations or wild crop relatives has been shown to be useful to improve crop yields (Gur and Zamir 2004), and has also been important for the genetic improvement of forest trees (Neale 2007). The extent of variability in the P. sibirica breeding populations was estimated by measuring population parameters including PCV and H2. In this study, PCV for H for the four provenances ranged from 27.87% (year 11) to 37.56% (year 18), and PCV for BD and CL also reached 37.60% and 31.14%, respectively, indicating wide variation among P. sibirica provenances. Within the same provenance, however, PCV for height gradually decreased with tree growth, suggesting the variation within a given provenance was smaller. The magnitude of H2indicated the reliability with which the phenotypic expression of a trait will reflect the genotype. In this study, the estimates of H2for H from year 11 to year 18 ranged from 0.967 to 0.983, and estimates of H2for BD and tree CL at year 18 were 0.974 and 0.968, respectively. High estimates for heritability indicated that trait selection can be effective, without being overly influenced by environmental effects (Chandrababu and Sharma 1999).
Determining trends in age-to-age correlations was relatively straightforward for growth and yield traits, requiring only patience and repeated assessment. Shortening the breeding cycle of trees through early selection can produce more genetic gain per year if there are strong genetic correlations between early and mature traits (Gon?alves 2005). There are many reported age-age correlations for growth in conifer species and rubber (Hevea brasiliensis) (King and Burdon 1991; Matheson et al. 1994; Gon?alves et al. 1998; Gon?alves 2005). In this study, there were strong correlations between H, DBH and CL at year 18 after introduction. The strong age-to-age correlations were also found for H in different years (11–18), and all correlation coefficients appeared to be significantly positive. This indicates that the selection for P. sibirica provenance was practical, and that yield predictions were feasible.
As a result of long-term natural selection, plant species and populations adapt to the range of environmental conditions that define their habitats. The performance of plants introduced to new regions always shows a strong dependence on the original environment. In this study, growth traits of the introduced Siberian pine provenances were markedly correlated with their source environments. After long term survival in the new environment, however, the influence of the source environment declined over time. The introduced trees adapted to the new environment gradually, and they displayed excellent growth, with no negative effects from plant diseases and insect pests. These results indicate a successful introduction of P. sibirica to northeast China. Site effects reflect the reaction of trees to the combined effects of edaphic and local and regional climatic conditions (Pliura et al. 2007). The North Hanggay provenance had lower survival, was more sensitive to climatic fluctuations, and possessed a lower wood biomass as well as CL, which could possibly reflect the obvious geographical differences between the source environment and the study site. The latitude of the experimental site was 51.43N, which was farther north than North Hanggay source site (47.48N). The elevation of the North Hanggay site was 2414 m, 2.4 times higher than the elevation at the experimental site (1000 m). The geographical parameters of the other three source sites were closer to those of the experimental site. From the climate data we can also see that the average annual temperature of North Hanggay was 3.4°C, but the average annual temperatures at the other locations were approximately 0°C. At the experimental site, the annual temperature averaged -4°C, which could possibly be another important factor leading to high mortality in the trees from North Hanggay.
Conclusions
The objectives justify the methods in tree breeding. In this study, considering growth traits and survival rates, the Novosibirsk, Tomsk, and Altai Mountain provenances of P. sibirica showed better performance than the North Hanggay provenance. But lessons learned from over 50 years of experience with Pinus radiata in Australia indicated that proper definition of breeding objectives, combined with adoption of modern selection and breeding methods for population improvement can result in progeny that show significant economic benefits in early cycles (Wu et al. 2007). Emerging approaches, such as genomic selection (Grattapaglia and Resende 2011) have potential to greatly accelerate breeding of forest trees. Although the apparent successful introduction of P. sibirica into China was an excellent example of introducing an exotic tree species, there were several shortcomings to the present study: (1) the seeds were collected in four P. sibirica habitats that represented only a small part of the natural geographic range and genetic variation of this species; (2) the study was conducted at a single test site with relatively small populations; and (3) traits related to reproduction (e.g. cone and seed production) were not measured. These subjects should be addressed in future studies.
We thank Professor Guangyi Zhao, Meijun Zhang and Shuyun Lin for growing the seedlings measured in this study.
Alantsev NK. 1981. The stone pine. Moscow: Forestry Industry Press, pp. 60–61.
Carrasquinho I, Gon?alves E. 2013. Genetic variability among Pinus pinea L. provenances for survival and growth traits in Portugal. Tree Genetics & Genomes, 9: 855–866.
Chandrababu RJ, Sharma RK. 1999. Heritability estimates in almond [Prunus dulcis (Miller) D.A. Webb]. Scientia Horticulturae, 79: 237-243.
Danusevicius J. 2001. Use of introduced provenances to increase genetic diversity in local scots pine populations. Biologija, 1: 59-61.
Engelmark O, Sjoberg K, Andersson B, Rosvall O, Agren G, Baker W, Barklund P, Bjorkman C, Despain D, Elfving B, Ennos R, Karlman M, Knecht M, Kinght D, Ledgard N, Lindelow A, Nilsson C, Peterken G, Sorlin S, Sykes M. 2001. Ecological effects and management aspects of an exotic tree species: the case of lodgepole pine in Sweden. Forest Ecol Manag, 141: 3-13
Grattapaglia D, Resende MD. 2011. Genomic selection in forest tree breeding. Tree Genetics and Genomes, 7: 241-255.
Gon?alves P, Bortoletto N, Cardinal A, Gouvea L, Costa R, Moraes M. 2005. Age-age correlation for early selection of rubber tree genotypes in Sao Paulo State, Brazil. Genet Mol Biol, 28: 758-764.
Gon?alves P, Bortoletto N, Fonseca F, Bataglia O, Ortolani A. 1998. Early selection for growth vigour in rubber tree genotypes in Northwestern Sao Paulo State (Brazil). Genet Mol Biol, 21: 515-521.
Gur A, Zamir D. 2004. Unused natural variation can lift yield barriers in plant breeding. PLoS Biol. 2(10) e245. Doi: 10.1371/journal.pbio.0020245
Hai PH, Jansson G, Harwood C, Hannrup B, Thinh H. 2008. Genetic variation in growth, stem straightness and branch thickness in clonal trials of Acacia auriculiformis at three contrasting sites in Vietnam. For Ecol Manag, 255: 156-167.
Hansen JK, Roulund H. 1996. Genetic parameters for spiral grain, stem form, Pilodyn and growth in 13 year old clones of Sitka Spruce (Picea sitchensis (Bong.) Carr.). Silvae Genet, 46: 107–113
Isaev AS. 1985. Stone pine forest in Siberian. The USSR academy of science, Novosibirsk. P. 30.
Iroshnikov AI, Politov DV. 2004. Five-needle pines in Russia: Introduction and breeding. USDA Forest Service Proceedings RMRS-P-32: 64–72.
King JN, Burdon RD. 1991. Time trends in inheritance and projected efficiencies of early selection in a large 17-year old progeny test of Pinus radiata. Can J For Res, 21: 1200–1207.
Krylov GV, Talantsev NK, Kozakova NF. 1983. The cedar [“Kedr”]. Moscow: Forestry Industry Press.73–34.
Li JY, Brunner AM, Meilan R, Strauss S. 2008. Stability of transgenes in trees: expression of two reporter genes in poplar over three field seasons. Tree Physiology, 29: 299–312.
Mack RN, Lonsdale WM. 2001. Humans as global plant dispersers: getting more than we bargained for. BioScience, 51: 95–102.
Matheson A, Spencer DJ, Magnussen D. 1994. Optimum age for selection in Pinus radiata using basal area under bark for age:age correlations. Silvae Gen, 43: 352–357.
Marron N, Ceulemans R. 2006. Genetic variation of leaf traits related to productivity in a Populus deltoides×Populus nigra family. Can J For Res, 36: 390–400.
Neale DB. 2007. Genomics to tree breeding and forest health. Current Opinion in Genetics and Development, 17: 539–544.
Peterken GF. 2001. Ecological effects of introduced tree species in Britain. For Ecol Manag, 141: 31–42.
Pliura A, Zhang SY, Mackay J, Bousquet J. 2007. Genotypic variation in wood density and growth traits of poplar hybrids at four clonal trials. For Ecol Manag, 238: 92–106.
Politov DV, Krutovsky KV. 2004. Phylogenetics, genogeography and hybridization of five-needle pines in Russia and neighboring countries. USDA Forest Service Proceedings RMRS-P-32, 85–97.
Wu HX, Eldridge KG, Matheson A C, Powell MB, Mcrae TA, Butcher TB, Johnson IG, Wu XH, Eldridge KG, Matheson AC, Powell MB. 2007. Achievements in forest tree improvement in Australia and New Zealand—Successful introduction and breeding of radiata pine in Australia. Australian forestry, 70(4): 215
Zhao XY, Ma KF, Shen YB, Zhang M, Li KY, Wu RL, Zhang ZY. 2012. Characteristic variation and selection of forepart hybrid clones of Sect. Populus. Journal of Beijing Forestry University, 34: 45–51.
 Journal of Forestry Research2014年3期
Journal of Forestry Research2014年3期
- Journal of Forestry Research的其它文章
- Effects of shifting cultivation on biological and biochemical characteristics of soil microorganisms in Khagrachari hill district, Bangladesh
- An integrated method for matching forest machinery and a weight-value adjustment
- Finite element analysis of stress and strain distributions in mortise and loose tenon furniture joints
- The in fl uence of silane coupling agent and poplar particles on the wettability, surface roughness, and hardness of UF-bonded wheat straw (Triticum aestivum L.)/poplar wood particleboard
- A modified Murashige and Skoog media for efficient multipleshoot induction in G. arborea Roxb.
- Characterization of expressed genes in the establishment of arbuscular mycorrhiza between Amorpha fruticosa and Glomus mosseae