
Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using Maxent modelling
2022-11-04 09:23:52DanLiuXiangdongLeiWenqiangGaoHongGuoYangshengXieLiyongFuYuancaiLeiYutangLiZhuoliZhangShouzhengTang
Journal of Forestry Research 2022年6期
Dan Liu · Xiangdong Lei · Wenqiang Gao · Hong Guo · Yangsheng Xie ·Liyong Fu · Yuancai Lei · Yutang Li · Zhuoli Zhang · Shouzheng Tang
Abstract Knowledge on the potential suitability of tree species to the site is very important for forest management planning. Natural forest distribution provides a good reference for aff orestation and forest restoration. In this study,we developed species distribution model (SDM) for 16 major tree species with 2,825 permanent sample plots with natural origin from Chinese National Forest Inventory data collected in Jilin Province using the Maxent model. Three types of environmental factors including bioclimate, soil and topography with a total of 33 variables were tested as the input. The values of area under the curve (AUC, one of the receiver operating characteristics of the Maxent model)in the training and test datasets were between 0.784 and 0.968, indicating that the prediction results were quite reliable. The environmental factors aff ecting the distribution of species were ranked in terms of their importance to the species distribution. Generally, the climatic factors had the greatest contribution, which included mean diurnal range, annual mean temperature, temperature annual range,and iosthermality. But the main environmental factors varied with tree species. Distribution suitability maps under current(1950 - 2000) and future climate scenarios (CCSM4-RCP 2.6 and RCP 6.0 during 2050) were produced for 16 major tree species in Jilin Province using the model developed. The predicted current and future ranges of habitat suitability of the 16 tree species are likely to be positively and negatively aff ected by future climate. Seven tree species were found to benef it from future climate including Betula costata, Fraxinus mandshurica, Juglans mandshurica, Phellodendron amurense, Populus ussuriensis, Quercus mongolica and Ulmus pumila; f ive tree species will experience decline in their suitable habitat including B. platyphylla, Tilia mongolica, Picea asperata, Pinus sylvestris, Pinus koraiensis;and four ( Salix koreensis, Abies fabri, Pinus densif lora and Larix olgensis) showed the inconsistency under RCP 2.6 and RCP 6.0 scenarios. The maps of the habitat suitability can be used as a basis for aff orestation and forest restoration in northeastern China. The SDMs could be a potential tool for forest management planning.
Keywords Species distribution model · National forest inventory data · Natural forest · Climate change · Site suitability mapping · Maxent modelling
Introduction
Understanding the potential suitability of tree species to the site is very important for forest management planning.Natural forest is the most stable forest ecosystems with the richest biodiversity and complex structure, where spatial information of distribution of tree species provides an important reference for matching tree species with the sites.The spatial distribution of tree species can be described by a species distribution model (SDM). The SDM mainly uses present data of the species distribution and environmental data to estimate the ecological niche of the species according to specif ic algorithms and ref lect the species preference to the habitats in the form of probability (Phillips et al. 2006).The results can be interpreted as the probability of occurrence of a species, habitat suitability or species richness.Therefore, SDM can be used as one of the important tools for forest management planning (Falk and Mellert 2011;Dolos et al. 2015; M?rkel and Dolos 2017).
The common species distribution methods mainly include the Climate Change Experiment (CLIMEX) (Lanoiselet et al. 2002) as the earliest one, the Bioclimate Analysis and Prediction System (BIOCLIM) (Busby 1991), Domain analysis (DOMAIN) (Carpenter et al. 1993) based on bioclimatic data, the Genetic Algorithm for Rule-set Production(GARP) (Stockwell 1999) and machine learning methods including the Maximum Entropy Species Prediction Model(Maxent) (Phillips et al. 2004) based on the ecological niche,ANN(artif icial neural network), regression trees and support vector machines (SVM) (Elith and Leathwick 2009).In recent years, Maxent has become one of the most widely used SDMs (Phillips et al. 2006; Liu et al. 2018; Feng et al.2020; Kaky et al. 2020; Li et al. 2020; Lin et al. 2020; Méndez-Encina et al. 2021; González-Hernández et al. 2020).The SDMs were widely used in space ecology, conservation planning, biological reserve designing, habitat restoration and management, plant community and ecosystem modeling(Zhang et al. 2016), and therefore it has become an eff ective tool (Raxworthy et al. 2003; Elith and Leathwick 2009;Butler et al. 2016; Cao et al. 2016). The SDMs were also applied in assessment of the impact of the species diversity pattern, prediction of the potential distribution of the endangered species, economically valuable species (Guisan et al.2006; Duan et al. 2014; Butler et al. 2016), environmental risk assessment, invasive species management (Cabeza et al. 2004; Li et al. 2014), and prediction of the eff ects of the global environmental change on the species and ecosystems (Iverson et al. 2004; Ballesteros-Barrera et al. 2007;Lobo et al. 2010; Gogol-Prokurat 2011; Benito-Garzón et al.2013). However, to our knowledge, SDM has been used in limited studies on forest management decisions for tree species suitability to sites (Fourcade et al. 2014; Gastón et al.2014).
The largest natural forest region in China is northeast forest with an existing forest area of 33.47 million ha,accounting for 13.86% of the total forest in the country. The stocking volume is 3.16 billion m 3 , accounting for 18.51%of the total forest in the country (State Forestry and Grassland Administration 2019), which plays an important role in timber production and ecological services. Most of the existing studies on natural forest restoration have focused on species introduction experiments, experiences and opinions of vegetation experts. At present, the study of species distribution in this forest region is conf ined to only a few tree species and forest types, such asQuercus mongolicaFischer ex Ledebour, Larch and mixed Korean pine-broadleaved forests (Wu 2003; Wang et al. 2006; Leng et al. 2008; Yin et al. 2013). A systematic evaluation on the suitability of the geographical distribution of major tree species in the natural forests of northeastern China are lacking, especially for some rare and high-value species, such asPinus koraiensisSiebold et Zuccarini,Phellodendron amurenseRupr.,Juglans mandshuricaMaxim., andFraxinus mandshuricaRupr.Species distribution, composition, and growth condition in natural forests are the important references for aff orestation and other silvicultural plan, thus, it is necessary to evaluate the suitability of the geographical distribution of tree species in natural forests. A set of the permanent sample plots of Chinese National Forest Inventory (NFI) provided a good database for this study, but so far, this database has not been used to explore the distribution and site suitability of various tree species. Therefore, this study aimed at (1)developing the SDMs for 16 major tree species based on the Chinese NFI data in Jilin Province, northeastern China;(2) investigating major environmental factors aff ecting the distribution of tree species and site suitability; (3) analyzing the potential distribution under current and future climate;and (4) generating site suitability maps for 16 major tree species in Jilin Province.
Materials and methods
Study area
The study area covers the regions of Jilin Province, northeastern China, and lies between 121°38’ - 131°19’ E and 40°52’ - 46°18’ N, which belongs to the temperate continental monsoon climate. From southeast to northwest, the climate changes from humid to semi-humid to semi-arid.The province-wide annual mean temperature ranges are 2 °C - 6 °C. The average annual temperature in the winter is below - 11 °C. The average temperature of the plain in the summer is above 23 °C. Generally, annual frost-free period are 100 - 160 d. The average number of sunshine hours in Jinlin Province are 2,259 - 3,016 h. The average annual precipitation is 400 - 600 mm, but seasonal and regional diff erences are large, with 80% precipitation in summer and most precipitation in the east. The study area includes about 2,000 m altitude gradient. The soil in the province mainly includes dark brown forest soil, black soil, chernozem soil and chestnut soil, among which the area of dark brown forest soil is the largest. Coniferous tree species includePinus koraiensis,Larix olgensisHenry,Picea asperataMast.,Abies nephrolepis(Trautv.) Maxim.,Pinus densif loraSieb.et Zucc. andPinus sylvestrisvar.mongolicaLitv.; the broadleaf tree species includeBeula emaniiCham.,Quercus mongolica,Betula platyphyllaSuk.,Fraxinus mandshuricaRupr.,Juglans mandshuricaMaxim.,Populus ussuriensisKom. andTilia amurensisRupr. (Forest of Jilin Province 1988).
Data and methods
Tree species dataThis study used data collected from the permanent sample plots of the 8 th NFI in Jilin Province. The NFI survey is based on the 4 km × 8 km grid system with a sample plot area of 600 m 2 . Among the total 8,872 NFI samples plots in the Province, only 2,825 sample plots in the natural forests were selected for our purposes. From the composition of the tree species, tree species appearing as frequently as possible and available records of sample plot locations, 16 major tree species with sample size more than 40 were selected with 13,869 observations. They represented major tree species in the region covering coniferous/deciduous and late-successional/pioneer ones. The sample sizes of each tree species are shown in Table 1. Plot distributions by tree species with natural origin were shown in Fig. S1.Environmental dataThe climate data including 19 bioclimatic variables (Table 2)were taken from the WorldClim database (Hijmans et al.2005) in the f inest resolution available with a grid of 30"(1 km × 1 km). The current (1950 - 2000) and future (2050)climate data were used to develop SDMs and simulate changes in habitat suitability for 16 tree species. Two future climate scenarios (CCSM4-RCP 2.6 and CCSM4-RCP 6.0)were selected. With a registration of the basic geographic data of Jilin Province, the bioclimatic data of the nearest grid to the center of the sample plot were used.
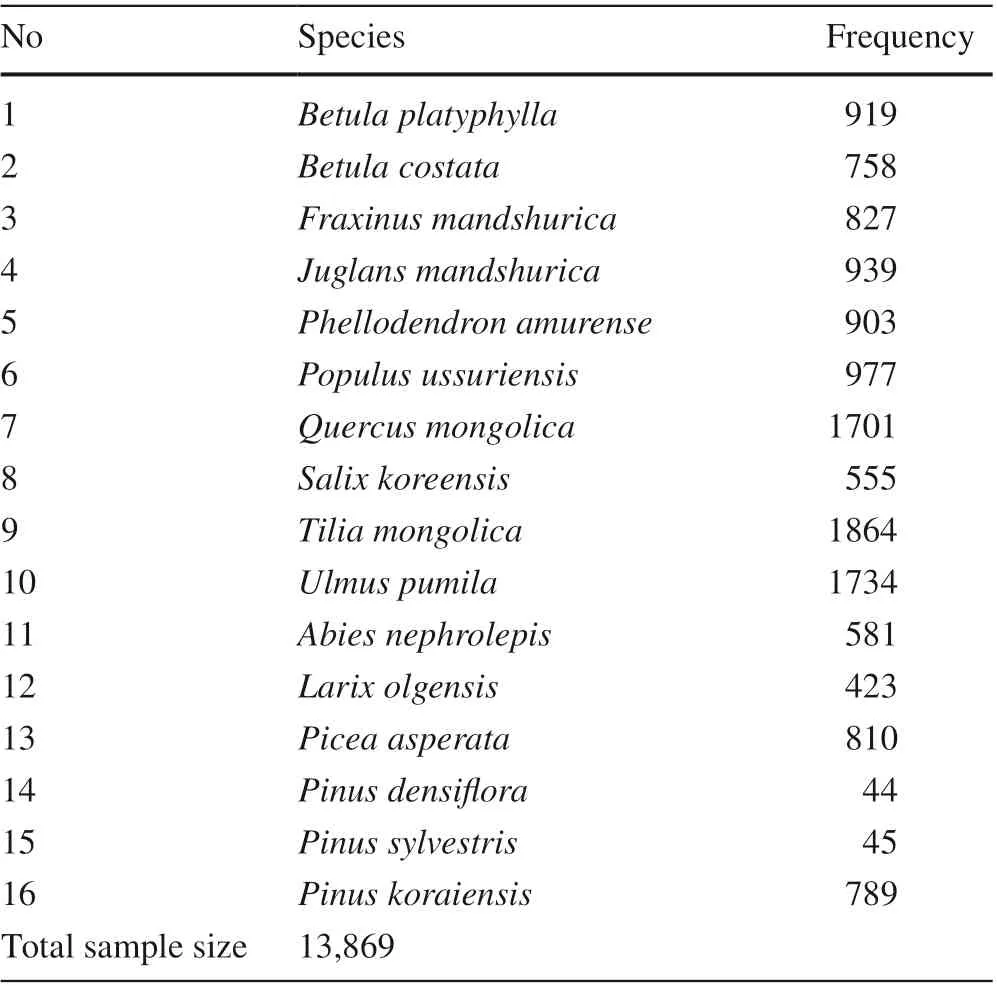
Table 1 Sample sizes of the main tree species of the natural forest in Jilin Province for species distribution simulation
Soil data were downloaded from SoilGrids system( https:// soilg rids. org ), which provides global predictions with 250-m resolution for standard numeric soil properties at seven standard depths (0, 5, 15, 30, 60, 100 and 200 cm)(Hengl et al. 2017). The basic def initions of these 11 factors are shown in Table S1. The soil samples from the f irst f ive layers were weighed and averaged as the attribute values of the soil in the corresponding layer. The combined layer was used as the soil variable input to the Maxent model in this study.
A 1: 4,000,000 vector map of China was downloaded from the National Geomatics Center of China ( http:// ngcc.sbsm. gov. cn , accessed on March 2016) as the base map for the analysis, and the scope of the study area (Jilin Province)was cropped. Based on the digital elevation model (DEM)data at a 30-m resolution in Jilin Province, the elevation,slope and aspect layers were generated, and the resolution and coordinates system were consistent with those in the environmental layers, such as climate and soil.
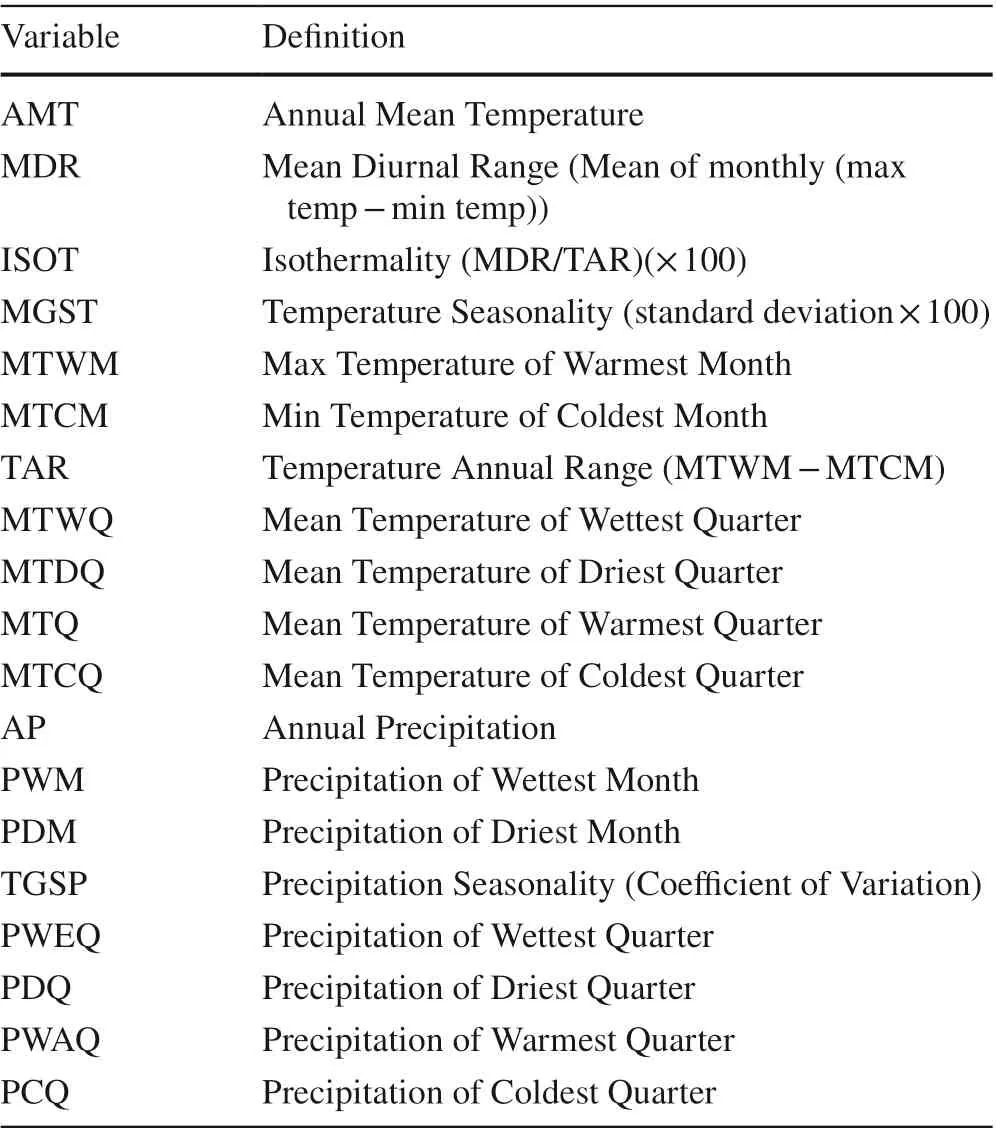
Table 2 Summary of bioclimatic variables used in this study
Environmental variable selection
To avoid multicollinearity problems, correlation analysis was f irstly explored among 33 candidate environmental variables (19 bioclimatic, 3 topographic, and 11 soil). Variables that were highly correlated (r> 0.7) were evaluated based on expertise and the importance from initial SDMs tests. Finally, only 14 variables were retained including eight bioclimatic variables (MDR, AMT, TAR, ISOT, PDQ, AP,MTDQ, MTWM), two topographic variables (aspect, slope)and four soil variables which are soil pH (pH), soil organic carbon content in the f ine earth fraction (soc), volumetric fraction of coarse fragments(> 2 mm) (cfvo), and bulk density of the f ine earth fraction (bdod).
Species distribution model
In this study, Maxent model was selected since it is one of the most widely used SDMs and showed good performance for species distribution modelling in the literature (Raxworthy et al. 2003; Cao et al. 2016; Elith and Leathwick 2009;Gogol-Prokurat 2011; Brown et al. 2017; Liu et al. 2018;Narouei-Khandan et al. 2020). As a SDM based on the maximum entropy theory, the distribution with the largest entropy was selected from the eligible distribution center as an optimal distribution. The dependent variables for the Maxent model were the present data of 16 tree species. There are 14 environmental variables as mentioned above. A total of 13,869 distribution sites was used as a sample for model operation. These present data and environmental variables were input into the Maxent software (Version 3.3.3 k) (Elith et al. 2015; Merow et al. 2013; Phillips et al. 2017), with the maximum iteration numbers of 5000 and the convergence threshold of 10 -5 . The gained value represented the distribution probability, whose value ranged between 0 and 1. With the gained value increasing, the probability of the presence of the tree species also increased. Finally, the distribution probability map of each tree species was generated under three climate scenarios (Current, RCP 2.6 and RCP 6.0). For the classif ication of the site suitability, based on the standard of possibility evaluation by the Intergovernmental Panel on Climate Change (Phillips et al. 2017), f ive levels of site suitability were def ined: extremely suitable, very suitable,suitable, very unsuitable, and extremely unsuitable sites.
Receiver operating characteristic (ROC) curves were created by plotting sensitivity vs. specif icity and ten-fold crossvalidation AUC (area under the curve) scores were used to evaluate the accuracy of the resulting model (Metz 1978,Phillips et al. 2004). The range of the AUC is [0, 1]. Models with an AUC scores of 0.5 indicate a model performing no better than random, while models with an AUC scores of 1 indicate a perfect model (Phillips et al. 2004; Phillips et al.2006; Elit hand Leathwick 2009; Elith et al. 2015). The larger value of the AUC indicates the better model performance and the more reliable the prediction results. According to the def inition provided by Philips (Phillips et al.2006), the accuracy is poor with AUC [0.5, 0.6], fair with AUC [0.6, 0.7], good with AUC [0.7, 0.8], very good with AUC [0.8, 0.9], and extremely good with AUC [0.9, 1.0].
The results of the constructed Maxent model were used to quantitatively evaluate the inf luence of the 14 factors (climate, soil and topographical factors) on the geographical distribution and site suitability of each tree species under three climate scenarios. The importance of the environmental variables was judged by the Jackknife method, which is a general calculation method with the assumption of a testing and conf idence interval used to obtain the score of each environmental factor in aff ecting the distribution of the specif ic tree species under current climate condition. Finally,the top four major environmental factors of each tree species were determined.
Results
The accuracy of Maxent model
The Maxent model was evaluated using the training and test samples, and the AUC values of the SDMs were obtained for the 16 major tree species in the natural forest of Jilin Province (Table 3). Among 16 tree species, two species showed extremely good, 11 showed very good, and 3 showed good in terms of accuracy. Therefore, the prediction results of the potential distribution of the 16 species in Jilin Province by the Maxent model were reliable.
AUC curves showed that the training and test samples for the model corresponding to most of the tree species (14 species) were consistent (not shown), but two tree species,Pinus sylvestrisandPinus densif lora, also showed a large diff erence. Therefore, in general, the constructed model can be used to determine the distribution suitability of the main tree species in Jilin Province.
Major factors aff ecting the distribution of the main tree species
According to the Jackknife test provided by the Maxent model, the obtained scores of the inf luence of each environmental factor on the geographic distributions of 16 tree species were used to determine its importance (Fig. 1 a and b).The length of each variable line represents the score of each variable; that is, the variable is more important followed bythe line lengthen. In the graph, the horizontal axis represents the score of the regularized training gain, and the vertical axis represents each environmental factor. Dark blue, light blue, and red represent the scores of the current factor alone,the sum of the scores of the other factors except for the current factor, and the sum of the scores of all the variables,respectively.
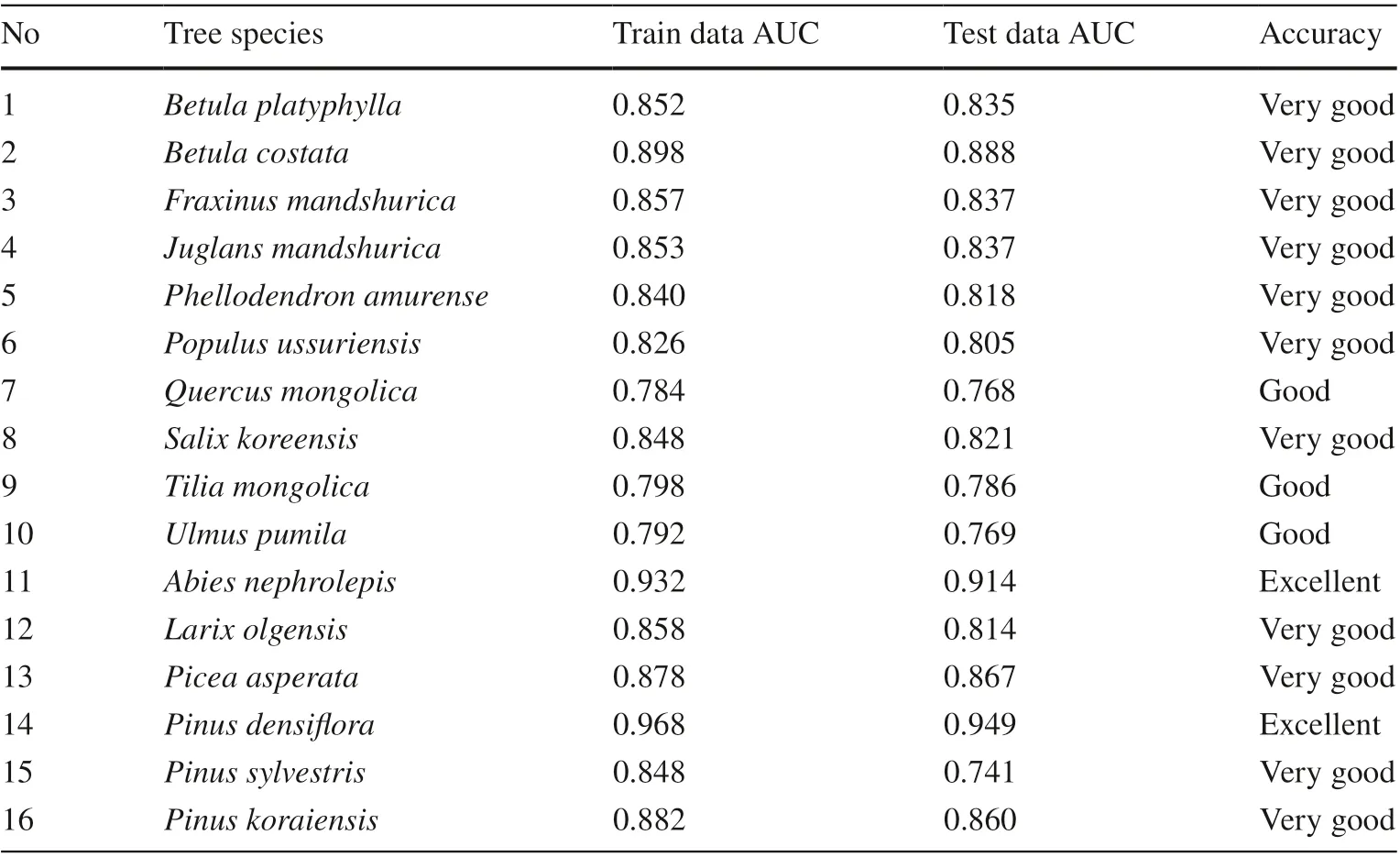
Table 3 The SDMs accuracy(AUC value) of the major tree species in Jilin Province
Accordingly, the main climatic factors aff ecting the distribution of the tree species included Mean Diurnal Range(MDR), Annual Mean Temperature (AMT), Temperature Annual Range (TAR) and Isothermality (ISOT); the soil factors are pH and soil organic carbon content in the f ine earth fraction (SOC) and the topographic factor is slope.The main environmental factors corresponding to diff erent species were substantially diff erent. Mean Diurnal Range(MDR) was common for all tree species. For the conifer tree species, onlyPinus densifl orawas closely related to Precipitation of Driest Quarter (PDQ) andPinus sylvestriswas closely related to annual precipitation (AP). All broadleaf tree species were closely related to pH, andFraxinus mandshurica,Juglans manshurica,Phellodendron amurense,Populus ussuriensis,Quercus mongolica,Salix koreensis,Tetraena mongolicaandUlmus pumilawere related to soil organic carbon content in the f ine earth fraction (SOC). The PDQ and AP only aff ected the adaptability ofPinus densif lora.The slope only aff ected the adaptability ofJuglans mandshuricaandPinus sylvestris. Among the top four environmental factors inf luencing the site suitability identif ied for each tree species (Table 4), MDR was listed for all tree species; pH for 14 trees species; SOC for 11 trees species;and AMT for 9 trees species.
Table 5 showed the percentage indicating the impact of climate, soil and topography on the geographic distribution and site suitability of each tree species. The values were calculated based on the accumulated environmental impact values of each tree species. The statistical analysis was performed after classifying the bioclimatic, soil and topographical factors. Results showed that climate was the most important variable, and the soil was the second, but topography was the least important. Soil and climate showed similar contribution to the distribution only forLarix olgensis.
Mapping the distribution suitability of 16 tree species under current and future climate
Tree species distribution suitability under current climate
According to the constructed SDMs, the grade of the suitability zone of each tree species in each suitability distribution area was obtained based on the bioclimatic, soil and topographic data of each kilometer grid (Fig. S2).
Generally, the distribution range of tree species decreased from southeast to northwest in the province. The distribution suitability varied signif icantly with the tree species. To make it clear, Fig. S3 gave the subregions of the Provinces for distribution suitability analysis. Among the six coniferous species,Pinus sylvestriswas widely distributed and found suitable for most areas in the eastern part and some small areas in the western part of Jilin Province. From the growth perspective, the medium growing coniferous species, such asPinus densifl ora,Larix olgensis,Pinus sylvestrisandPinus densif lorhad the smallest area for its suitability. The high suitable zone forAbies fabriwas fragmented and concentrated, and its suitable areas in the eastern part was also not continuous.Larix olgensis’s extremely high and high suitable zones were very narrow, but there were large area ofcontinuous low suitable areas, very suitable to distinguish in a small part of Antu, Fusong, Changbai counties (Fig. S3).
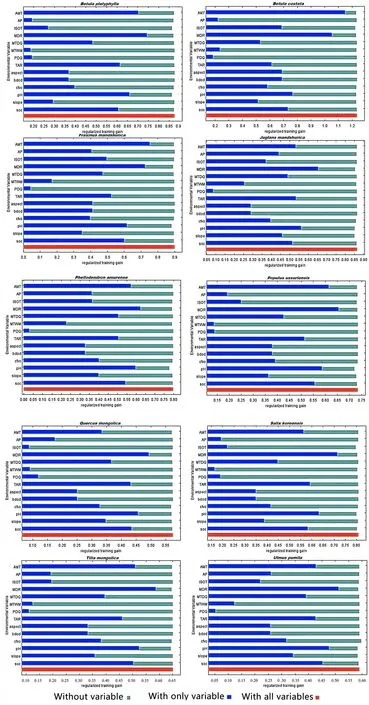
Fig. 1 Jackknife training gain of each environmental variable for the broadleaf and conifer tree species
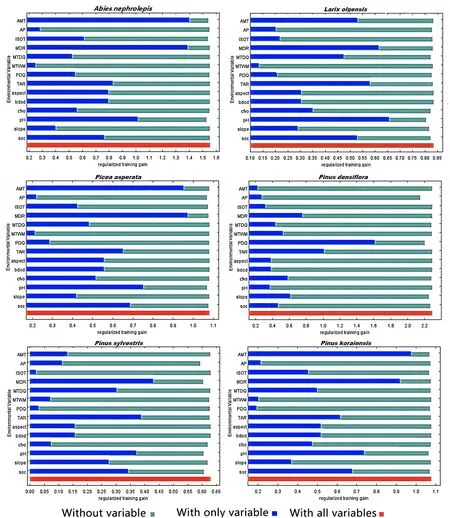
Fig. 1 (continued)
Slow growing coniferous species, such asPicea asperataandPinus koraiensis, both had large high suitable area in Changbai Mountains. Four fast-mid growing broad-leaved tree species, includingPopulus ussuriensis,Salix koreensis,Betula platyphyllaandUlmus pumila, the suitable areas were mainly concentrated in the hilly area of the east mountain of the Great Black Hills. Both the high and extremely high suitable zones of theUlmus pumilawere the largest,followed byBetula platyphylla,Populus ussuriensisandSalix koreensis. The suitable area forBetula costatawas smaller and not continuous.Tetraena mongolicahad a large and continuous high suitable zone in the Dongning and Wangqing. In contrast,Betula costatain high suitable fracture area was not coherent.
Quercus mongolicaas common broadleaf species in northeast China, was continuous high suitable in the eastern part of Jilin Province (Hunchun and Dongning). In the western Taonan area there was also a small low suitability area.
Fraxinu mandshurica,Juglans mandshurica, andPhellodendron amurensewere the three major valuable hardwood tree species in northeastern China.Fraxinu mandshuricawas one of the ancient relict floraspecies originated from the tertiary period, which mainly appeared in the eastern mountain to northeastern China and had an important scientific value for the study of the flora in the Paleogene and the climate in quaternary ice age. The distribution also showed these species were suited to eastern region with high ranks.
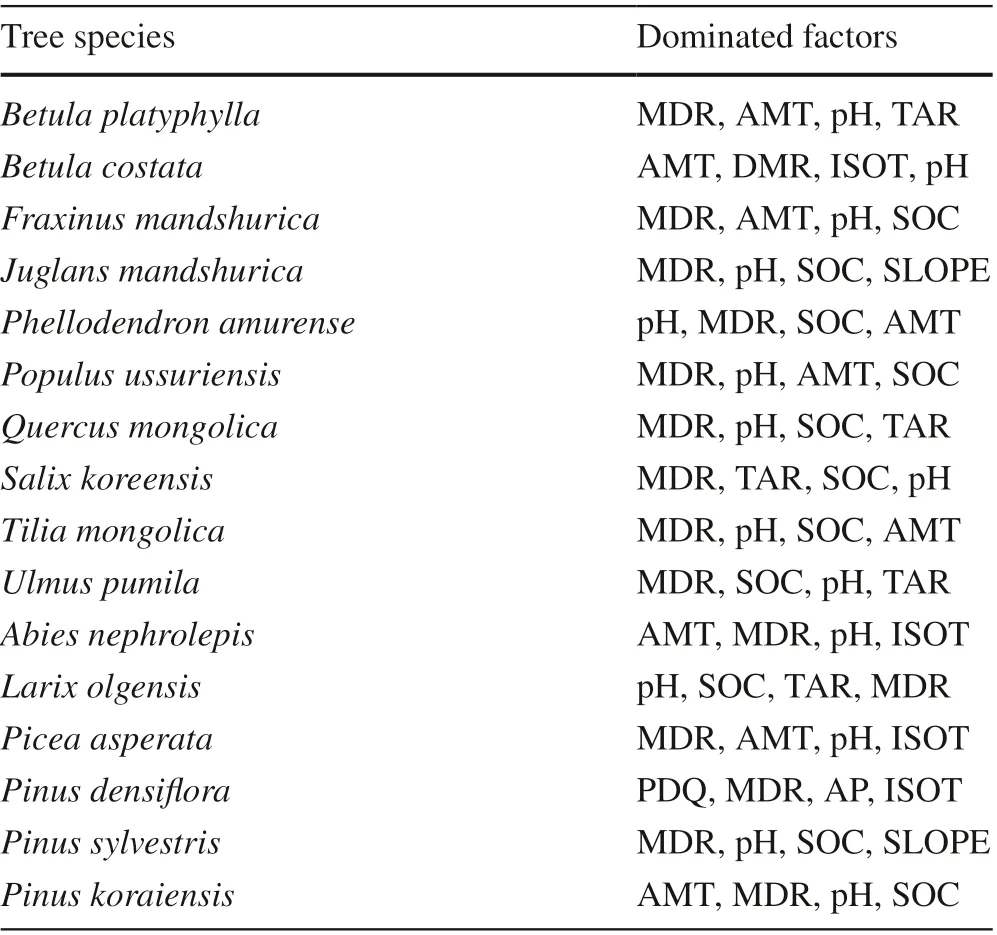
Table 4 The top four dominant factors aff ecting the geographic distribution of various tree species in natural forests
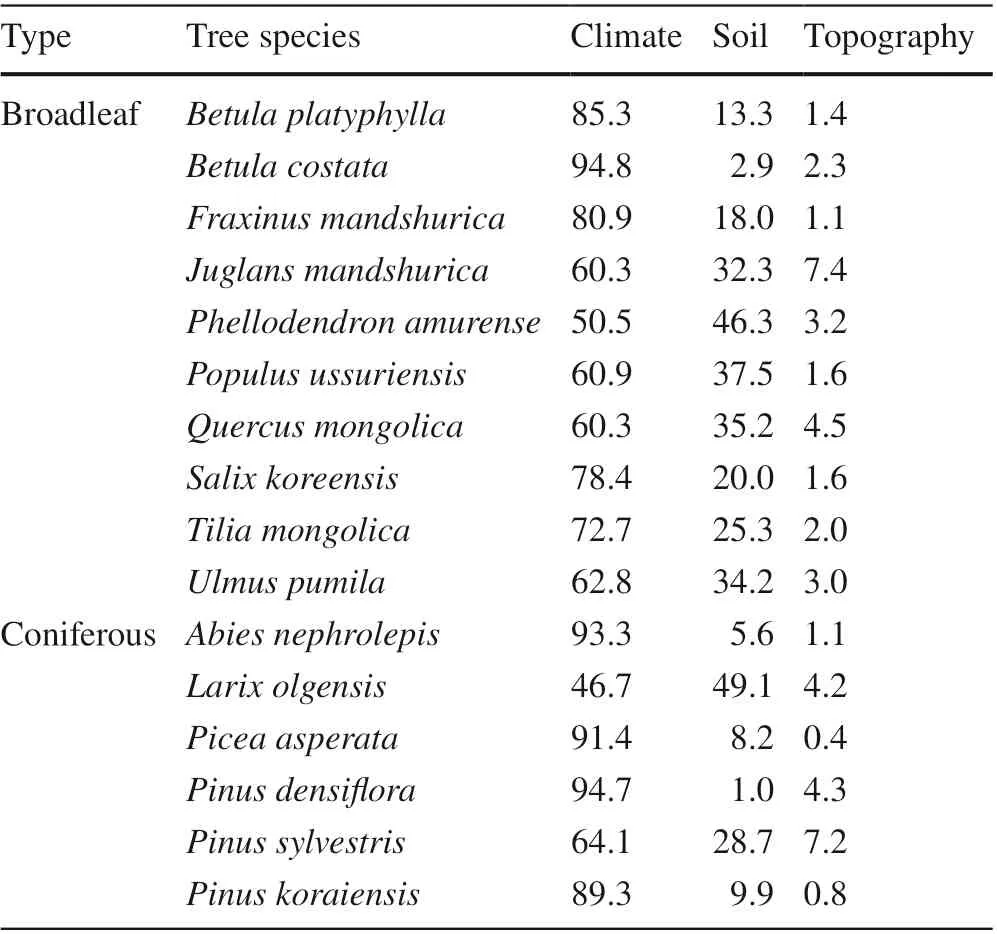
Table 5 Contributions of various environmental variables to the distribution suitability of each tree species
Tree species habitat suitability under future climate change
According to the results of the Maxent modelling, the predicted current and future ranges of habitat suitability of the 16 tree species are likely to be positively and negatively aff ected by future climate (Fig. 2). Seven tree species were found to benef it from future climate includingBetula costata,Fraxinu mandshurica,Juglans mandshurica,Phellodendron amurense,Populus ussuriensis,Quercus mongolicaandUlmus pumila; f ive tree species will experience decline in their suitable habitat includingBetula platyphylla,Tetraena mongolica,Picea asperata,Pinus sylvestris,Pinus koraiensis; and four (Salix koreensis,Abies fabri,Pinus densif loraandLarix olgensis) showed the inconsistency under RCP2.6 and RCP6.0 scenarios. The percent changes in suitable (extremely suitable, very suitable, suitable) and unsuitable habitat (very unsuitable and extremely unsuitable) ranged - 2.23% to 2.29% and - 4.11% to 8.73%,respectively.
Discussion
Maxent model performance
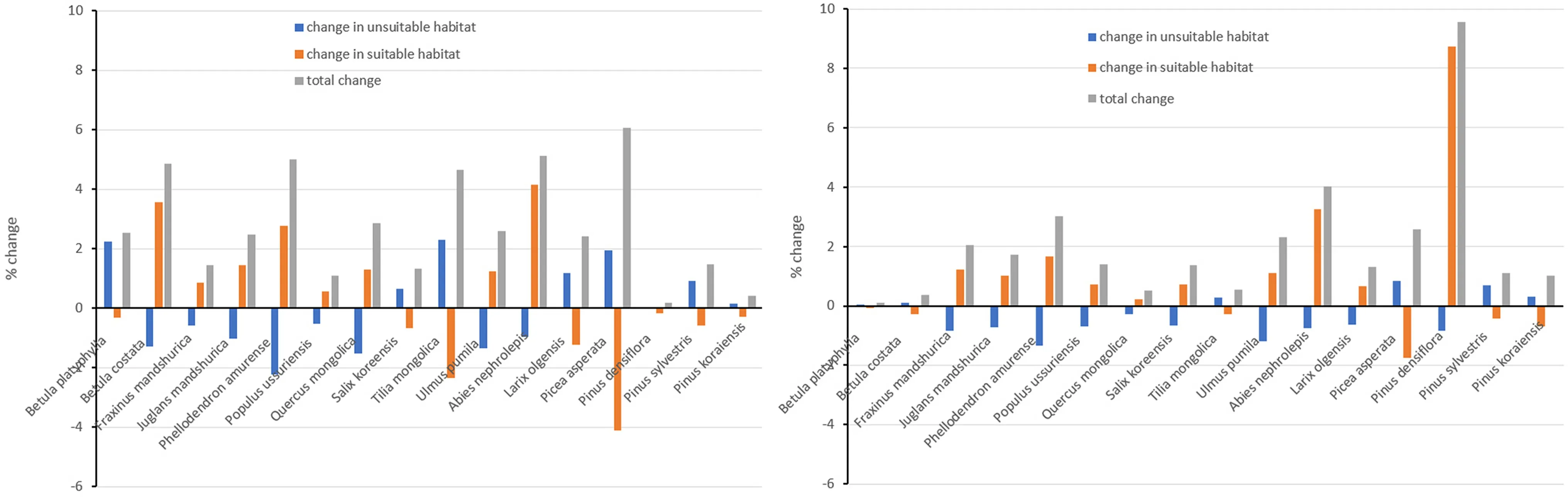
Fig. 2 Percent change in habitat suitability of the 16 tree species under current and projected climate scenarios
The SDM is mainly based on the known distribution of the species and its environmental variables to investigate the niche and potential distribution of the species. Among existing methods, Maxent is a machine-learning method that uses an iterative approach to approximate species distributions based on the principles of maximum entropy, in which habitat suitability is predicted across the landscape based on the values of environmental variables at known occurrence locations (Raxworthy et al. 2003; Phillips et al.2006; Gogol-Prokurat 2011). We applied Maxent to model the distribution of 16 major tree species in Jilin Province of northeast China based on 33 environmental variables related to bioclimate, soil and topographic under three climate conditions, and to determine the main environmental variables of each species that aff ect the distribution suitability. The AUC values of training and test dataset for the distributions of 16 tree species ranged from 0.784 to 0.968.Among of which, two reached the level of “extremely good accuracy”, eleven reached “very good accuracy”, and three reached “good accuracy”. However, the trained and tested samples showed a large diff erence forPinus densif loraandPinus sylvestris, which is related to the small sample size(Zhang et al. 2016; Li et al. 2020), and should be careful for generalization. These implied that our models had excellent predictive ability, and the distribution of 16 tree species was therefore well described by the selected predictors. The results indicated that the prediction accuracy of the developed Maxent model in this study is very high and acceptable with good generalization ability. Although AUC is widely used as the standard method to assess the accuracy of predictive distribution models, there was a debate on its use (Lobo et al. 2008; Jiménez-Valverde 2012). Lobo et al.( 2008) pointed that AUC was a misleading measure of the performance because AUC ignores the predicted probability values and the goodness-of-f it of the model, assumes equal costs for commission and omission errors, and is sensitive and positively related to the spatial extent under consideration. Measures deriving from a contingency table and the ROC curve were considered as more informative than reporting just a single AUC value (Jiménez-Valverde 2012). In this study, AUC values together ROC curves were applied for model assessment. The Maxent model has been shown to outperform other methods and has consistently high accuracy even when limited species occurrence data is available (Raxworthy et al. 2003; Hernandez et al. 2006;Gogol-Prokurat 2011; Zhang et al. 2016). For example,Gogol-Prokurat ( 2011) predicted the habitat suitability for rare plants at local spatial scales in EI Dorado County, California in USA using a species distribution model based on the Maxent model. They found that the developed Maxent produced models were successful at discriminating between suitable and unsuitable habitat at the local scale for all four species, and predicted habitat suitability values were proportional to likelihood of occurrence or population abundance for three of four species. Zhang et al. ( 2016) modeled distribution of 6828 woody plants using Maxent and removed the collection bias using null model in China. Yin et al. ( 2013) also used this model to study the distribution ofQuercus mongolicaand obtained the AUC of 0.923. The advantages of the Maxent model are: (a) It is a model of the maximum information entropy in all the models that satisfy the constraints. As a classic model, it has higher accuracy rate although this is debatable; (b) The constraints can be set f lexibly, and the degree of adaptation of the model to the unknown data and the degree of f itting of the known data can be adjusted by how much the constraint conditions are (Kaky et al. 2020). Recently, the Maxent software has announced a new open-source release which further facilitate the work in improving the science of modeling species distributions(Phillips et al. 2017). In addition, ensemble models combining predictions from single SDM models theoretically should produce more accurate and robust predictions than a single model, and showed the signif icant improvement of the model accuracy (Araújo and New 2007; Grenouillet et al.2011). We did not test other SDMs and ensemble models in the study, but multiple model comparisons and fusion will be examined for better prediction of tree species distribution in the future study.
Environmental factors aff ecting the distribution of the tree species
The distribution of tree species is aff ected by a combination of various biotic and abiotic factors, such as the environment and competition, and their geographic distributions are profoundly aff ected by climate change. The results of this study showed that among the climate, soil and topographical factors, bioclimatic factors had the greatest impact on the distribution of tree species. Climate at the regional scale is the most important factor that determines the type and distribution of vegetation on the earth (Lin et al. 2020;Zhang 1993), which was also supported by previous study in this region (Leng et al. 2008). The predictors related to temperature were the main factors aff ecting habitat suitability for most tree species although the specif ic variables diff ered which included MDR, AMT, TAR and ISOT. In particular,MDR and MAT were the most important climatic predictors. They can directly regulate the photosynthesis process and further aff ect plant production and reproduction through the intensity of the light (Hawkins et al. 2003). The results were partly consistent with previous studies in the region.Yin et al. ( 2013) found the dominant factors aff ecting the distribution ofQuercus mongolicain China were annual precipitation, annual mean temperature, accumulated temperature greater than 5 °C and highest monthly temperature. Jia et al. ( 2017) reported that the dominant factors were annual precipitation, precipitation seasonality, elevation, annual mean temperature, and mean temperature of wettest quarter for mixed broadleaved-Korean pine forests (includingPinus koraiensis,Quercus mongolica,Tetraena mongolicaandFraxinus mandshurica). The inf luencing factors in our study were annual mean temperature, mean diurnal range and temperature annual range for these four tree species.Leng et al. ( 2008 ) investigated three types of larch and found that the climatic factors inf luencing the distribution ofL.lanceolataincluded average annual precipitation, precipitation in the growing season and average temperature in January; in this study, they are mean diurnal range and temperature annual range. In addition, soil pH and soil organic carbon also aff ect tree species distribution, especiallyLarix olgensis.Soil pH has an enormous inf luence on soil biological, chemical, and physical properties and processes that aff ect plant presence and growth. Soil organic carbon (SOC)is crucial in maintaining soil quality because of its role in improving soil nutrition. The reason on the diff erence in bioclimatic variables may be related to the data sources and the distribution of samples. For example, Yin et al. ( 2013)and Jia et al. ( 2017) used data from the Chinese Virtual Herbarium, the Teaching Specimen Resource Sharing Platform and the National Specimen Information Infrastructure. NFI data were selected in our study with systematic layout and full coverage. In addition to environmental competitions,pests and diseases, forest gaps and extreme weather can contribute to the disappearance of a tree species and hence aff ect the distribution. This study did not consider the impacts of these factors. Biotic interactions also inf luence the ranges and abundances of species, which are the new challenges for species distribution modelling (Zhan 1995; Elith et al.2006; Anderson 2017).
The habitat suitability mapping and its implications for forest management
Based on SDMs, maps of the habitat suitability of the 16 species under current and future climate was generated at 1 km × 1 km resolution across Jilin Province, which can be used accordingly as a basis for aff orestation and forest restoration in northeastern China. We can get the latitude and longitude of each forest type in the study area and the counties and cities where the core suitable areas are located. It could be in accordance with the division of sub regions by forest site in China, where Jilin is divided into three site sub regions(Fig. S3) including 3B (Eastern Songliao plain), 3C (Western Songliao plain) and 7B (central Changbai Mountain) (Zhan 1995), from the perspective of site suitability rank analysis of each natural tree species.
Owing to the importance of climate in habitat suitability modelling showed in the study, future habitat suitability maps were produced under RCP2.6 and RCP6.0 scenarios. Both positive and negative eff ects of climate change on habitat suitability were found for 16 tree species. Among of them,late-successional tree species includingFraxinus mandshurica,Juglans mandshurica,Phellodendron amurense,Quercus mongolicashowed the increase of habitat suitability, butPinus koraiensisandPicea asperatashowed the decrease. For poineer tree species,Betula platyphyllaandPinus sylvestrisshowed negative, butPopulus ussuriensisandUlmus pumilapostitve;Larix olgensisandPinus densif lorashowed both positive and negative under RCP2.6 and RCP6.0, respectively.Dyderski et al. ( 2018) reported that pioneer and coniferous species was more threatened than mid- and late-successional species because of their better drought tolerance. Our results were partly consistent with these f indings. Although late-successional tree speciesPinus koraiensispresented as negative to climate change, the change was very small (less than 0.3%).Generally, the climate change in northeast China showed the rising temperature and the increased precipitation. Our models presented that the temperature-related factors demonstrated a greater contribution, which indicates higher sensitivity of tree species to temperature changes. Diff erent pine species have diff erent sensitivity to temperature, and thus the diff erence in habitat suitability. Species distribution modeling generates valuable information for sustainable forest management, making policy decisions and conservation management for natural and planted forests in northeastern China. Based on the results,we should pay special attention to tree species with negative eff ects of future climate change, and take measures such as adjusting and transforming species compositional structure.For tree species in suitable distribution areas, we should focus on strengthening forest tending to develop its potential productivity. Our f indings can be applied in various ways such as the identif ication of additional localities where these tree species may already exist, but has not yet been detected; the recognition of localities where it is likely to spread to; the priority selection area for introduction and cultivation and the conservation management of such rare tree species. Furthermore, main types of the forests will be examined for adaptive management in the future study, especially the natural distribution of mixed forests and the impact of future climate change on their distribution.
Conclusions
Based on the permanent sample plots with natural origin from Chinese NFI data collected in Jilin Province, this study for the f irst time constructed the SDMs for 16 major tree species in northeastern China using the Maxent model. The results showed a clear discrimination among the site suitability to all 16 major tree species. The AUC values of training and test datasets were between 0.784 and 0.968, indicating that the prediction results were quite reliable. Temperaturerelated climate variables were the most important variable for the potential distribution of most tree species. Maps of habitat suitability under current and future climate were produced for 16 tree species in Jilin Province using the Maxent model. Tree species showed diff erent response to future climate change, both negative and positive eff ects were found.These maps can be used as a basis for aff orestation and forest restoration in Jilin Province. Our results also showed the potential of SDM as a basic tool for forest management planning.
Acknowledgements We are grateful to Ram P. Sharma for his valuable suggestions on this study
 Journal of Forestry Research2022年6期
Journal of Forestry Research2022年6期
- Journal of Forestry Research的其它文章
- Mixing planting proportions in a plantation aff ects functional traits and biomass allocation of Cunninghamia lanceolata and Phoebe bournei seedlings
- Intraspecif ic leaf morphological variation in Quercus dentata Thunb.: a comparison of traditional and geometric morphometric methods, a pilot study
- Correction to: Eff ects of active molecules of Korean pine seed on rodent health and implications for forest regeneration
- Seed, germination, and seed-reserve traits diff er along an altitudinal gradient
- Analysis of the chloroplast genomes of four Pinus species in Northeast China: Insights into hybrid speciation and identif ication of DNA molecular markers
- Genetic variation and superior provenances selection for wood properties of Larix olgensis at four trials