
Photosynthetic acclimation to long-term high temperature and soil drought stress in two spruce species(Picea crassifolia and P.wilsonii)used for afforestation
2018-03-19 05:08:12XiaoweiZhangLitongChenJingruWangMinghaoWangShuliYangChangmingZhao
Journal of Forestry Research 2018年2期
Xiaowei Zhang?Litong Chen?Jingru Wang?Minghao Wang?Shuli Yang?Changming Zhao
Introduction
Water availability and temperature are considered to be the main variables limiting photosynthesis,affecting growth and survival of plants(Niu et al.2008;Ghannoum and Way 2011;Gago et al.2013).Photosynthesis is sensitive to environmental variables which will be profoundly affected by future climate change,including elevated air temperatures and decreased water availability(Luo 2007;Gunderson et al.2010;Lin et al.2012;Ashraf and Harris 2013).Meanwhile,the ability of plants to modify photosynthesis in response to high temperatures and/or drought stress has been shown to be species-speci fi c resulting from different photosynthetic acclimation potentials(Way and Oren 2010;Ashraf and Harris 2013;Way and Yamori 2014).Therefore,the potential for photosynthetic acclimation to new growing conditions plays a central role in the effects of climate change on plant growth and survival(Smith and Dukes 2013;Sendall et al.2015).
Photosynthetic acclimation can alter the short-term abiotic factor-response functions of photosynthesis associated with maintaining leaf gas exchanges under different growing conditions(Smith and Dukes 2013;Zhang et al.2015;Aspinwall et al.2016).For example,thermal acclimation of photosynthesis hasresulted in a shift in optimum temperature(Topt)and/or a change in sensitivity,revealed,for example,by a change in the shape of the temperature response curves(see Berry and Bjo rkman 1980;Hikosaka et al.2006;Way and Yamori 2014;Sendall et al.2015).Furthermore,the potential for plants to maintain photosynthetic capacity when wateravailability decreases depends on the sensitivity of leaf gas exchange to drought(Limousin et al.2013).However,plants have at least three different strategies for maintaining photosynthesis in response to changing temperatures or water availability.First,photosynthetic capacity is closely linked to stomatal conductance(Cond)(Flexas and Medrano 2002;Hikosaka et al.2006;German and Roberto 2013).Long-term elevated air temperatures or drought may alter the sensitivity of stomatal apertures,thus limiting photosynthesis(Zhang et al.2001;Reddy et al.2004;Gao et al.2009;Greer and Weedon 2012).Second,elevated temperatures or drought may lead to changes in leaf anatomy and density,for example,changing the leaf mass per area(LMA)to affect the mesophyll conductance of CO2(Poorter et al.2009;Yamori et al.2009;Vasseur et al.2012;Heroult et al.2013;Drake et al.2015).These two strategies could in fl uence water loss and constrain CO2exchange from leaves.The third,photosynthesis also be limited through biochemical processes which was temperature and water availability dependent(Way and Sage 2008;Lin et al.2012;Limousin et al.2013;Aspinwall et al.2016).In fact,the biochemical processes may be related to photosynthetic attributes such as the amount and/or ef fi ciency of enzymes and photosystem II(PSII)activity.Nitrogen content per mass(Nmass)is an important feature of photosynthetic apparatus,indicative of the relative proportion of enzymes in photosynthetic processes(Yamori et al.2009;Aspinwall et al.2016).In addition,the maximum quantum yield of PSII(Fv/Fm)is a reliable diagnostic indicator of photosynthetic activity(Reddy et al.2004;Gao et al.2009;Ma et al.2010;Wang et al.2014).
Picea crassifoliaKom.andP.wilsoniiMast.are two endemic species in Western China which often form a dominant component in coniferous forests(Farjón 2001;Fu et al.1999;Zhao et al.2008).Due to their signi fi cant commercial and ecological value,P.crassifoliaandP.wilsoniiare widely used for afforestation in Northern China.For example,in Qinghai Province alone,there are 14,000 ha of planted spruce forest,accounting for~5.4‰of the total spruce forest areas(Han et al.2015).However,current climate models suggest increasing temperatures(raising about 10°C by 2100)and droughts in these regions(Zou et al.2005;IPCC 2013).Coniferous forests may be particularly sensitive to climate change,which may result in changes in carbon exchange and a serious threat to survival(Zhang et al.2015;Aspinwall et al.2016;Kroner and Way 2016).Therefore,the objectives of our study were to examine the effects of long-term high temperatures and drought on photosynthesis of these two important conifers and to determine which was likely to be of more bene fi t to future forest stability in Northern China.
Materials and methods
Plant materials and growing conditions
Seeds of each species were collected within their natural range(Picea crassifoliaKom:30.3–37.8°N,126.5–130.5°E,Alt.:2400–3600 m above sea level(a.s.l.);P.wilsoniiMast:33.7–40.8°N,101.6–116.8°E,Alt.:1400–2800 m a.s.l.).In 2005,seeds were germinated and grown indoors at Yuzhong campus,Lanzhou University(35°56′37′N,104°09′05′E,Alt.:1750 m a.s.l.Temperatures ranged during the growing season from 7.7 to 25.8°C)for one year;the seedlings received ample water and light.Seedlings were then transplanted into 24 cm(upper diameter)×16 cm(basal diameter)× 17 cm(depth)pots fi lled with a homogeneous mixture consisting of equal volumes of peat and perlite.One pot was designed three seedlings.All pots were periodically watered to fi eld capacity(FC)according to Ma et al.(2010).On June 15,2009(dayt1),25 pots of each species of uniform growth(c.20 cm tallP.crassifoliaand c.25 cm tallP.wilsonii)were selected and divided into two groups:one for the water stress experiment,the other for the high temperature experiment.Control pots( fi ve of each species)were used for both experimentsandgrownat25/15°C and80% FC conditions.
Experiment 1:Drought experiment
Each fi ve pots of each species were randomly divided into low[80% of maximum fi eld capacity(FC)],mild(60%FC),moderate(40%FC)and severe(20%FC)water stress treatments.Water stress levels continued until October 16,2009,and maintained at these levels by weighing the pots every two days.Pots were assigned a random position in an arti fi cial intelligent greenhouse with growth temperatures controlled at 25/15°C day/night[moderate temperature(MT)]by a temperature control system.All seedlings were grown under 12 h photoperiods with light levels of 300–400 μmol photon m-2s-1at seedling height by artifi cial light sources for automatic control.Unfortunately,allP.wilsoniiseedlings grown at 20%FC died during the experimental period.Seedlings of both species in the remaining 35 pots were alive at the end of this experiment(October 15,2009;dayt2).
Experiment 2:High temperature experiment
The remaining pots( fi ve pots)of each species were placed in another greenhouse with growth temperatures controlled at 35/25°C[high temperature(HT)].All seedlings were raised under 12 h photoperiods with light levels of 300–400 μmol photon m-2s-1at seedling height.In both greenhouses,the CO2concentration was maintained at~380 μmol mol-1and relative humidity at 50 ± 5%.Seedlings in the high temperature treatment were watered suf fi ciently to avoid any effects caused by extreme water de fi cit.The experimental period continued until October 15,2009(dayt2).
Gas exchange and chlorophyll fl uorescence measurements
Leaf-level gas exchange(Pn,CondandTr)measurements were made with a portable open-path gas exchange system and a conifer chamber(Li-6400 and 6400-07,LI-COR Biosciences,Inc.)on fully expanded current year-old twigs for each experiment.Three to fi ve seedlings per treatment and species were randomly selected between 10:00–13:30 h during sunny weather,when the temperatures were 25 or 35°C,depending on each growth conditions. During photosynthetic measurements, CO2concentration was maintained at 380 μmol mol-1using portable CO2/air mixture tanks with output controlled by a LI-6400-01 CO2injector(LI-COR Biosciences,Inc.).Light levels were maintained at approximately 800 μmol m-2-s-1(saturated light level)at measurement height provided by an external light source.In addition,photosynthetic temperature curves of seedlings in each temperature treatment at 5 °C intervals from 40 to 15 °C were recorded.To ensure that the whole plant was exposed to the desired temperature settings,temperatures were controlled by changing air temperatures of the growth chamber,and micro-changing with the Li-6400 temperature control model.When measurements at one temperature were complete,thechambertemperatureconditionswere adjusted.Seedlings were allowed to equilibrate to chamber conditions for a minimum of 30 min before measuring the same twigs again.All measurements were completed in one day.After measurements had been taken,needles were cut and leaf areas were determined with a LI-3000A portable area meter(LI-COR Biosciences,Inc.)to calculate gas exchange parameters on an area basis.The measured needles were dried at 65°C for 72 h and dry mass determined(LM;Precisa XT120A,Precisa Instruments.Ltd.,Switzerland).TheseLMandLAvalues were used to calculate leaf mass per area(LMA).
When gas exchange measurements were taken,the maximum quantum yield ofphotosystem II(PSII)was measured for leaves adapted to dark conditions during an acclimatization period of 30 min.Chlorophyll fl uorescencemeasurementswere taken with a portable pulse amplitude modulated fl uorometer FMS-2(Hansatech,King’s Lynn,Norfolk,UK).At least fi ve replicates from each treatment were taken.
Leaf nitrogen measurements
We measured the concentration of nitrogen(N)using samples used for gas exchange measurements.Dried samples were fi nely ground with a mortar and pestle,and sent to the Analytical Testing Center,Lanzhou University for analysis using a CHN analyzer(Vario EL,Elementar,Germany).
Modeling of photosynthetic temperature curves
Photosynthesis data from temperature response curves were used to determine temperature-dependence and fi tted to the following quadratic equation(Gunderson et al.2010;Niu et al.2008;Sendall et al.2015):

wherePTrepresents the mean net photosynthetic rate at temperatureTin°C;andPoptis the photosynthetic rate at the optimum temperature(Topt).Parameterbdescribes the spread of the parabola(Battaglia et al.1996).For a givenAoptandTopt,parameterbis smaller and the photosynthetic temperature parabola ‘broader’if photosynthesis is less sensitive to short-term temperature changes.
Long-term sensitivity of photosynthesis to temperature
We also calculated an index to quantify the degree of thermal acclimation ofPnin response to HT(Way and Oren 2010)based on the following:

The index of photosynthesis(Acclimpn)for each species was equal to the ratio ofPnin HT leaves(35°C)to MT leaves(25°C).As an index for the degree of acclimation,ifAcclimpnis close to 1.0,this indicates that temperature acclimation exhibited is high(Way and Oren 2010;Yamori et al.2009).
Statistical analysis
Quadratic fi tting was used to estimate temperature response functions for photosynthetic rates(15–40 °C).The experiment was arranged in a completely randomized design with 3–5 replicates.Differences in all traits were determined by analysis of variance(one-way and General Linear Model,Proc GLM)and Tukey’s test for multiple comparisons.All data are presented as mean±SE.Differences were considered signi fi cant atp<0.05.Statistical analysis was performed using SPSS 16.0(SPSS Inc.,Chicago,IL,USA).
Results
Effects of long-term drought stress on photosynthesis
As available soil water decreased,Pn,Cond,andTrdecreased signi fi cantly in both species(Table 1).In the low water treatment,Pn,Cond,andTrinP.wilsoniiwere higher than inP.crassifolia,particularly in the case of the latter two variables(p<0.05,Table 1).In contrast,P.crassifoliahad signi fi cantly higher values ofPn,CondandTrunder mild and moderate water conditions,leading to a relatively greater decrease ofPn,CondandTrinP.wilsoniiwith increasing water stress(Table 1).Furthermore,there were signi fi cant interactions between species and water treatments forPn,CondandTr,suggesting that photosynthetic response to water stress was different in the two species(Table 2).
The maximum quantum yield of photosystem II(Fv/Fm)was not signi fi cantly different betweenP.crassifoliaandP.wilsoniiunder the low water treatment.However,as drought stress increased,Fv/FmforP.crassifoliaonly decreased signi fi cantly in the moderate treatment,whileFv/FmforP.wilsoniidecreased signi fi cantly in both mild and medium treatments.Meanwhile,the value ofFv/FminP.wilsoniiwas lower than inP.crassifoliafor mild and moderate treatments(Fig.1;Table 2).Hence,P.wilsoniialso appeared more sensitive to drought stress for this character.
Compared with the low water treatment,increasing soil water stress generated a signi fi cant reduction in the value ofNmassfor bothPiceaspecies,with the exception ofP.wilsoniiin the mild treatment.Meanwhile,the variations inNmassbetween species were only apparent in the low water treatment(Fig.2).Therefore,Nmassdiffered,depending on both the watering treatments and species(Table 2).However,LMAfor both species was not signi fi cantly changed in either treatment(data not shown),while the values ofLMAforP.crassifoliawere obviously larger than those forP.wilsonii(see Table 3).
Effects of long-term high temperatures on photosynthesis
We designed the second experiment to test the effects of temperature as a major limiting factor.As in the low water treatment above,Pn,CondandTrinP.wilsoniimeasured at 25°C(MT)were signi fi cantly higher than inP.crassifolia.After long-term 35°C(HT)treatment,PnandCondforP.crassifoliawere signi fi cantly higher than forP.wilsonii,whereasTrmeasured forP.crassifoliawas signi fi cantly lowerwithconsequencesforarelativelygreaterreductioninPnforP.wilsonii(Fig.3).Hence,the value ofAcclimpnforP.crassifoliawas about 0.75,higher than forP.wilsonii(Fig.4).Meanwhile,there were signi fi cant differences in temperature,species and their interaction(Table 4).
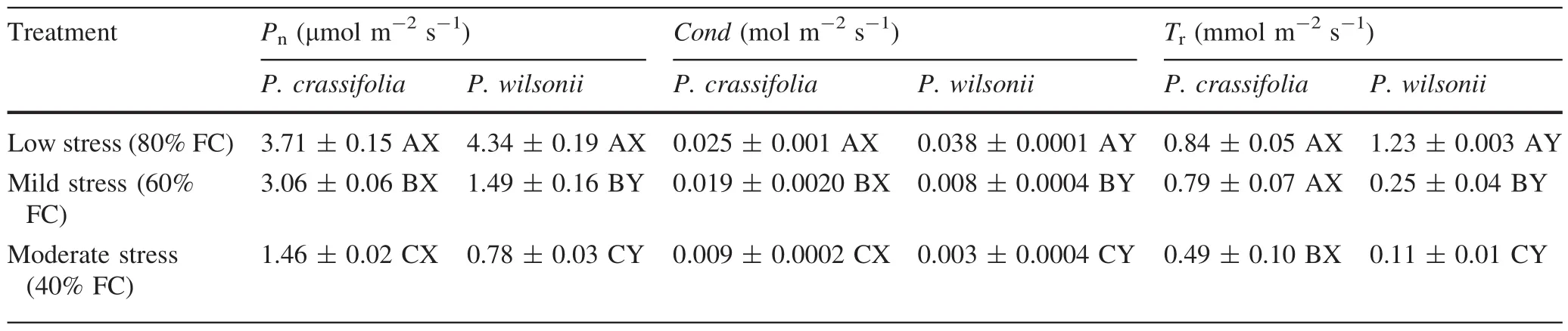
Table 1 Comparison of gas exchange parameters between Picea crassifolia and P.wilsonii,across different soil water treatments(80% of maximal fi eld capacity(FC),60 and 40%FC).Each value represents a mean and SE.Letters after SE values distinguish between statistically different(p<0.05)values for different water treatments(A,B,C)and between different species(X,Y)
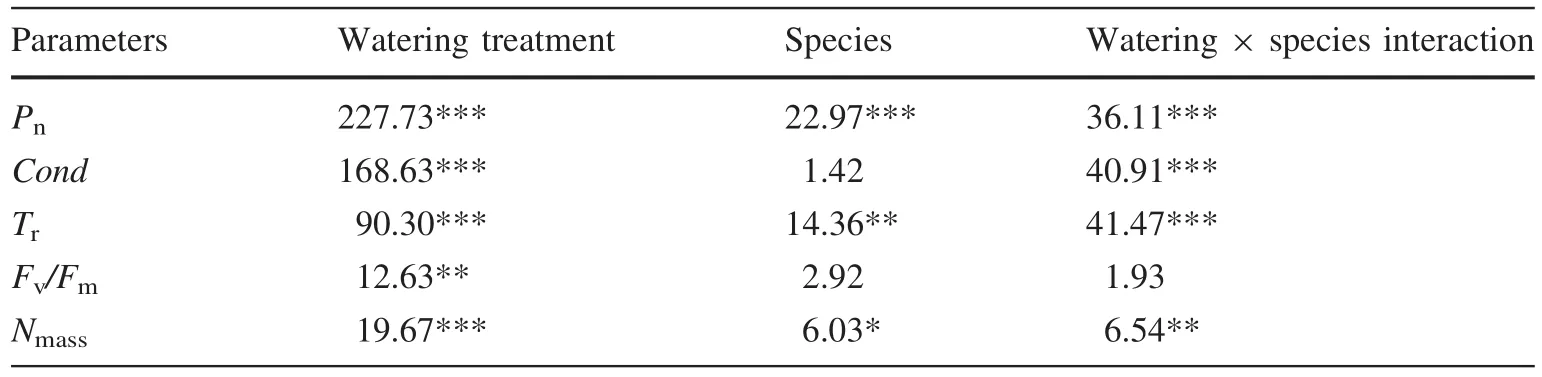
Table 2 Effects of watering treatment,species and their interaction on fi ve indicators measured during drought treatments
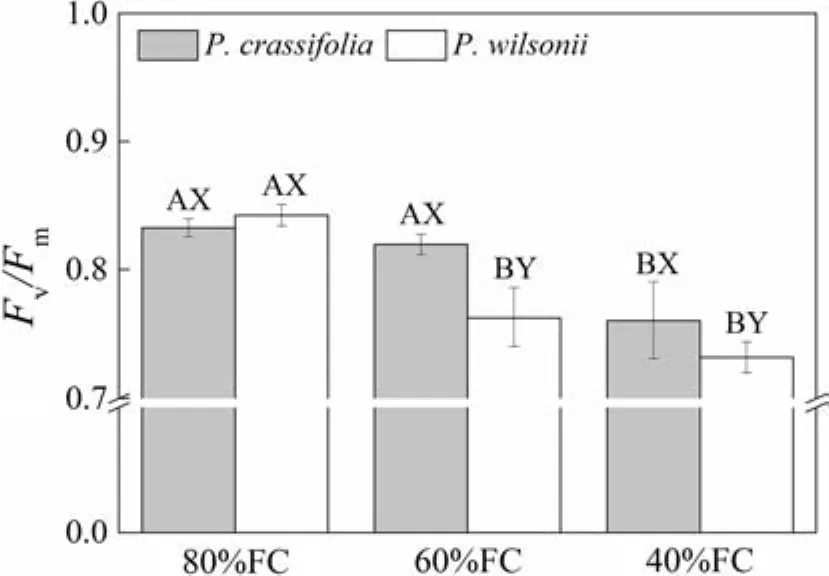
Fig.1 Maximum quantum yield of PSII(Fv/Fm)in seedlings of Picea crassifolia and P.wilsonii under different soil water conditions(80% of maximal fi eld capacity(FC),60 and 40%FC).Each value represents a mean and SE.Letters after SE values distinguish between statistically different(p<0.05)values for different water treatments(A,B)and between different species(X,Y)
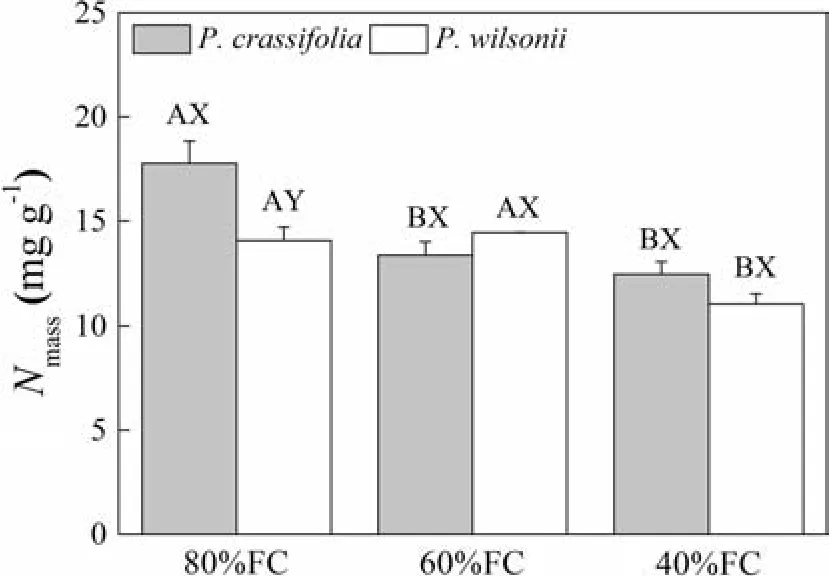
Fig.2 Effects of watering treatments on the nitrogen content per dry mass(Nmass)in seedlings of Picea crassifolia and P.wilsonii under different soil water conditions(80% of maximal fi eld capacity(FC),60 and 40%FC).Each value represents a mean and SE.Letters after SE values distinguish between statistically different(p<0.05)values for different water treatments(A,B)and between different species(X,Y)
At 25°C(MT),all estimated parameters(b,ToptandAopt)of the photosynthetic temperature response curves inP.wilsoniiwere signi fi cantly higher than those forP.crassifolia(Table 3).Following the 35°C(HT)treatment,the shapes of the curves were obviously different between species (Fig.5).OnlyToptforP.crassifoliawas signi fi cantly greater,whilstbandAoptwere reduced forP.wilsonii.Meanwhile,Aoptat 35°C inP.crassifoliawas higher than inP.wilsonii;however,there were no significant differences inbandToptbetween species(Table 3).Interactions between temperature and species for these variables were also highly signi fi cant,indicating that temperature treatments inb,ToptandAoptwere different betweenP.crassifoliaandP.wilsonii(Table 4).
For growth at 25°C(MT),Fv/Fmwas not signi fi cantly different between the two species.In contrast,Fv/FminP.wilsoniiwas signi fi cantly lower following the 35°C(HT)treatment for 4 months,and its value was clearly less than that forP.crassifolia(Table 3).Variations inLMAandNmasswere species-speci fi c following the 25°C(MT)treatment;LMAandNmasswere higher inP.crassifolia(Table 3).In contrast,there were no signi fi cant differences inLMAandNmassbetween species grown at 35°C(HT)(Table 3).However,there were only signi fi cant species effects with respect toLMA,while the values ofNmasswere signi fi cantly affected by temperature treatments(Table 4).
Discussion
Precipitation and temperature are the most important factors affecting plant growth and distribution because of their in fl uence on photosynthesis(Zhang et al.2009;Way and Oren 2010).In this study,we examined a combination of photosynthetic parameters and leaf morphological characteristics ofP.crassifoliaandP.wilsoniigrown under two temperature conditions and four water supply regimes.We found different patterns in their long-term response to temperature and drought:P.crassifoliaexhibited greater photosynthetic acclimation in both treatments as compared withP.wilsonii.
Photosynthetic acclimation to drought
Drought tolerance is essential for the survival and growth of many plants(Reddy et al.2004;Mao and Wang 2011).PnforP.crassifoliaandP.wilsoniidecreased with increasing drought stress(Table 1),suggesting that drought was inhibiting photosynthetic activity(Ashraf and Harris2013).Similar patterns have been observed in other plants(Ma et al.2010).However,photosynthetic responses to drought were different between the twoPiceaspecies in this study:the decrease ofPnforP.wilsoniiwas much larger than that forP.crassifolia,in mild and moderate water treatments,particularly in the mild water treatment(c.–65%;Table 1),suggesting that the photosynthesis ofP.wilsoniiwas more prone to drought limitations,whilst the photosynthesis ofP.crassifoliashowed an acclimatory response to drought.In addition,whilst the seedlings ofP.crassifoliasurvived the high stress treatment with water supplied at 20%FC,P.wilsoniiseedlings did not(seeMaterials and methods).This also suggests that high drought stress limits the growth and survival ofP.wilsonii,whileP.crassifoliais more resistant to drought,especially under extreme water de fi cit conditions.
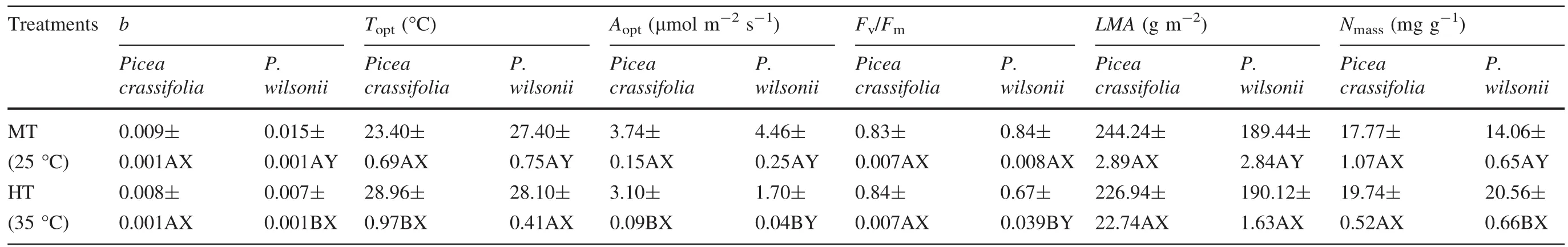
Table 3 Effects of growth temperature on several parameters (b,Topt,Aopt,Fv/Fm,LMA and Nmass) for Picea crassifolia and P.wilsonii
The higherPnunder drought observed inP.crassifoliacould be explained by the conditions affecting two processes.First,water supply was reduced with increasing drought stress,progressively inducing stomatal closure(Flexas and Medrano 2002;Reddy et al.2004;Gao et al.2009;Matteo et al.2014).Therefore,photosynthetic reduction in bothPiceaspecies may be partly explained by stomatal limitation.However,P.crassifoliahad higherCondandTrthanP.wilsoniiin the mild and moderate treatments.Moreover,larger decreases inCondofP.wilsonii(decreasingc.80%)were observed in the mild water treatment relative to non-stressed seedlings.These results suggest that the leaves of eachPiceaspecies showed different sensitivities to water de fi cit(Ashraf and Harris 2013),and that leaves ofP.wilsoniiwere more sensitive to drought.Second,with increasing water stress,drought can affect biochemical processes(Way and Sage 2008;Lin et al.2012;Limousin et al.2013).Our results reveal that decreases inFv/FmforP.crassifoliawere smaller in each treatment.There was only a slight reduction under the mild water treatment(Fig.1),suggesting more stable light capture and utilization inP.crassifolialeaves(Evans 1989;Reddy et al.2004).However,there was a signi fi cant decrease inNmassin the mild water treatment.In contrast,larger deceases inFv/Fmwere observed inP.wilsonii,suggesting that increasing water stress may inhibit or damage photosynthetic process inP.wilsonii(Ma et al.2010;Ashraf and Harris 2013).At the same time,steady values ofNmassinP.wilsoniiin the mild water treatment did not compensate for a larger reduction inFv/Fm.Thus,variations inFv/Fmalso partly explain different photosynthetic acclimations to drought in bothPiceaspecies.These results suggest that the photosynthetic process ofP.crassifoliahas a higher drought tolerance than that ofP.wilsonii.
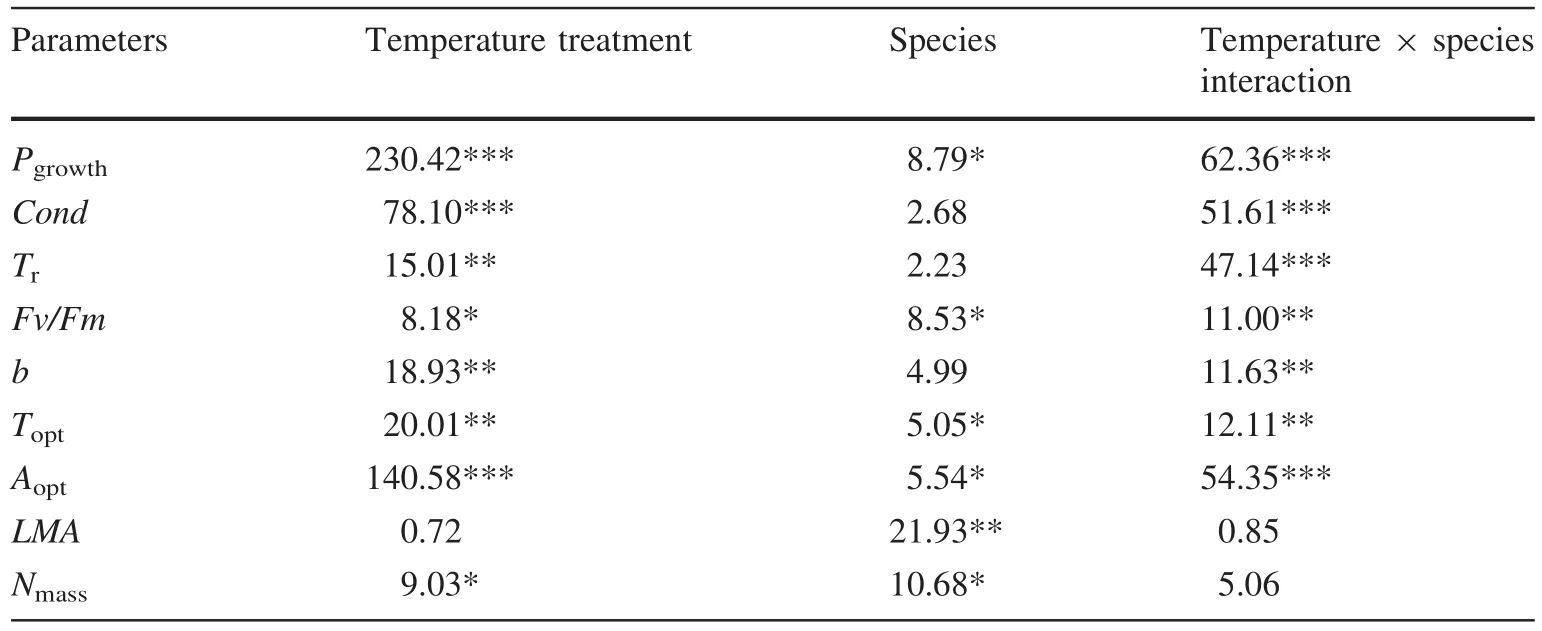
Table 4 Effects of growth temperature,species and their interaction on 10 measured indicators under the two temperature treatments
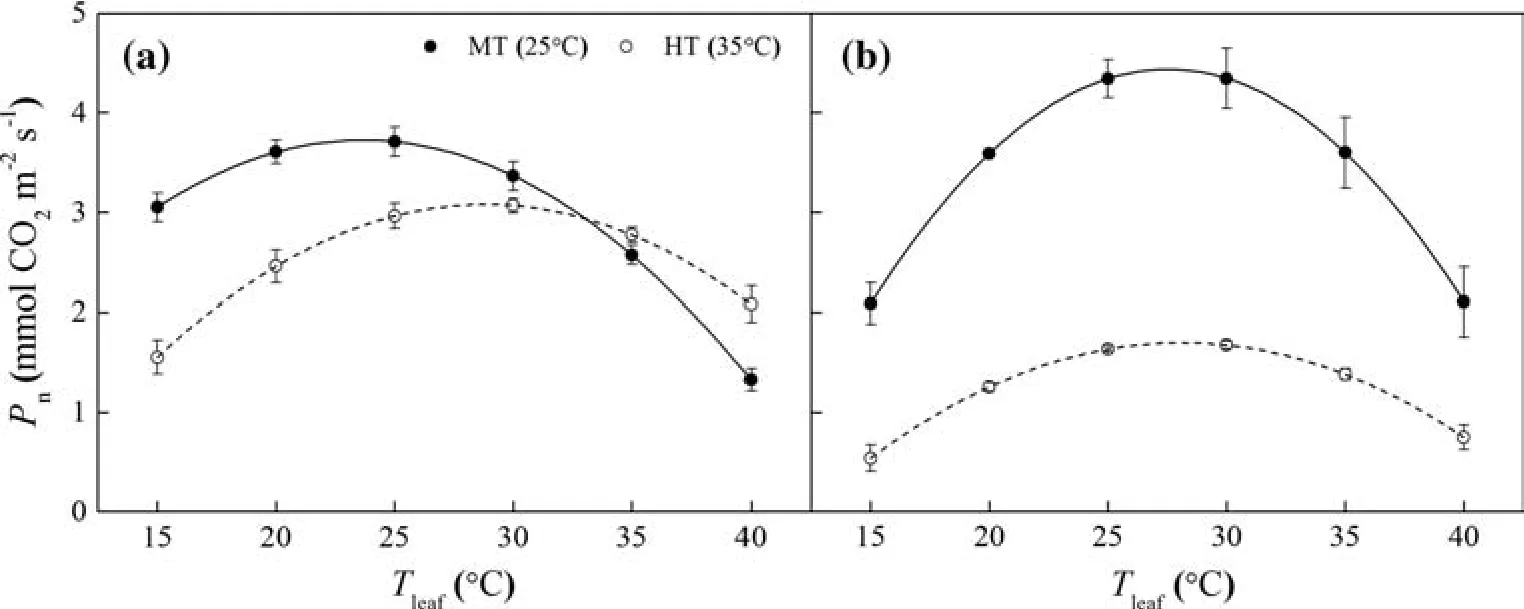
Fig.5 Temperature(Tleaf)responses of leaf photosynthetic rates(Pn)in Picea crassifolia(a)and P.wilsonii(b)under the MT and HT treatments.Data are presented as mean±SE
Photosynthetic acclimation to high temperatures
Grown at elevated temperatures,Pnmeasured at 35°C decreased in bothPiceaspecies compared to seedlings grown at 25°C,indicating that photosynthesis of these twoPiceaspecies did not completely acclimatize to elevated temperatures(Berry and Bjo rkman 1980).However,theAcclimPnforP.crassifoliawas higher than that forP.wilsonii(Fig.4),suggesting that thermal acclimation of photosynthesis inP.crassifoliais more effective than inP.wilsonii(Yamori et al.2009;Way and Oren 2010).P.crassifoliahas an inherently lower photosynthetic sensitivity to short-term temperature fl uctuations(e.g.parameterbin Table 3 and Fig.5;see Battaglia et al.1996;Niu et al.2008;Gunderson et al.2010;Sendall et al.2015).Photosynthesis ofP.wilsoniiwas more sensitive to short-term temperature fl uctuations(Table 3;Fig.5)and therefore the thermal sensitivity of photosynthesis inP.wilsoniiwas limited.Meanwhile,there was an upward shift in theToptofP.crassifoliathat reduced high temperature stress;this was not observed inP.wilsonii(Gunderson et al.2010;Way and Yamori 2014;Sendall et al.2015).Therefore,the photosynthetic process ofP.crassifoliahas a higher thermal tolerance than that ofP.wilsonii.
The regulation ofPnis related to changing stomatal conductance,leaf development,and biochemical processes during prolonged exposure to high temperatures(Way and Sage 2008;Lin et al.2012;Heroult et al.2013).Changes in stomatal conductance(Cond)could reducePnirrespective of biochemical effects(Hamerlynck and Knapp 1996;Zhang et al.2001;Hikosaka et al.2006;Greer and Weedon 2012).High temperatures are associated with increasing leaf-to-air vapor pressures leading to increasing drought stress.Potentially plants could limitCondto reduceTr(Day 2000).Our results show that decreasedCondforP.wilsoniilimitedTrat elevated temperatures(Fig.3),indicating increased heat-induced physiological drought stress forP.wilsoniieven though water de fi cits were avoided by providing abundant water during the experimental period(German and Roberto 2013).In contrast,increasedTrinP.crassifoliameant thatP.crassifoliaplants were exposed to thermal stress as a result of increasing water loss from leaves even when there was abundant soil moisture(Fig.3c).Hence,variations inCondbetween temperature treatments were important in explaining treatment differences inPn(German and Roberto 2013).On the other hand,LMAwas identical between temperature treatments for the two species(Tables 3,4),suggesting that high temperatures did not affect leaf structure and density.Leaf developmental processes are therefore unlikely to explain temperature treatment differences inPn.In addition,biochemical processes may partly explain the smaller proportional decreases inPnat high temperatures.Our results showed thatFv/Fmwas only signi fi cantly reduced by high temperatures inP.wilsonii,indicating that high temperature inhibits or damages the photosynthetic apparatus inP.wilsoniileaves.Hence,increasingNmassinP.wilsoniimay be assumed to repair chlorophyll and thylakoids(Evans 1989;Hikosaka et al.2006;Hikosaka and Shigeno 2009;Wang et al.2014).This was not compensated for by greater photosynthetic range.These results suggest that thermal acclimation of photosynthesis inP.crassifoliais more effective than inP.wilsonii(Fig.4).
Conclusions
Overall,our results suggest that stress caused by drought and high temperatures reduce thePnof the seedlings of bothPiceaspecies.However,P.crassifoliaexhibited higherphotosyntheticacclimation to both increasing drought and temperature thanP.wilsonii.In addition,we recorded higherCondandFv/FmforP.crassifoliawith increasing drought and temperature,indicating that these were responsible for the improved acclimation compared toP.wilsonii.Moreover,the photosynthetic apparatus inP.crassifolialeaves exhibited an inherently lower temperature-sensitivity and higher thermostability(see parameterb).Further,severe drought stress(20%FC)killedP.wilsonii.In conclusion,our results indicate thatP.wilsoniiis more susceptible to drought and high temperatures;P.crassifoliais more appropriate to plant to survive future climate increases and to sequester carbon.
AcknowledgementsWe thank Dr.David Blackwell for correcting the English in the fi nal manuscript.
Ashraf M,Harris PJC(2013)Photosynthesis under stressful environments:an overview.Photosynthetica 51(2):163–190
Aspinwall MJ,Drake JE,Campany C,Va?rhammar A,Ghannoum O,Tissue DT,Reich PB,Tjoelker MG(2016)Convergent acclimation of leaf photosynthesis and respiration to prevailing ambient temperatures under current and warmer climates in Eucalyptus tereticornis.New Phytol.doi:10.1111/nph.14035
Battaglia M,Beadle C,Loughhead S(1996)Photosynthetic temperature responses ofEucalyptus globulusandEucalyptus nitens.Tree Physiol 16:81–89
Berry J,Bjo rkman O(1980)Photosynthetic response and adaptation to temperature in higher plants.Ann Rev Plant Physiol 31:491–543
Day ME(2000)In fl uence of temperature and leaf-to-air vapor pressure de fi cit on net photosynthesis and stomatal conductance in red spruce(Picea rubens).Tree Physiol 20:57–63
Drake JE,Aspinwall MJ,Pfautsch S,Rymer PD,Reich PB,Smith RA,Crous K,Tissue DT,Ghannoum O,Tjoelker MG(2015)The capacity to cope with climate warming declines from temperate to tropical latitudes in two widely distributed Eucalyptus species.Global Change Bio 21:459–472
Evans JR(1989)Photosynthesis and nitrogen relationships in leaves of C3plants.Oecologia 78:9–19
Farjón A(2001)World checklist and bibliography of conifers.Royal Botanic Gardens,Kew,London,p 107
Flexas J,Medrano H(2002)Drought-inhibition of photosynthesis in C3plants:stomatal and non-stomatal limitations revisited.Ann Bot 89:183–189
Fu L,Li N,Mill RR(1999)Picea.In:Wu ZY,Raven PH(eds)Flora of China.Science Press and Missouri Botanical Garden Press,Beijing,pp 25–32
Gago J,Coopman RE,Cabrera HM,Hermida C,Molins A,Conesa MA,Galmes J,Ribas-Carbo M,Flexas J(2013)Photosynthesis limitations in three fern species.Physiol Plant 149:599–611
Gao DH,Gao Q,Xu HY,Ma F,Zhao CM,Liu JQ(2009)Physiological responses to gradual drought stress in the diploid hybridPinus densataand its two parental species.Trees 23:717–728
German VG,Roberto ACS(2013)Photosynthetic responses to temperature of two tropical rainforest tree species from Costa Rica.Tree 27(5):1261–1270
Ghannoum O,Way DA(2011)On the role of ecological adaptation and geographic distribution in the response of trees to climate change.Tree Physiol 3:1273–1276
Greer DH,Weedon MM(2012)Modelling photosynthetic responses to temperature of grapevine(Vitis viniferacv.Semillon)leaves on vinesgrown in a hotclimate.PlantCellEnviron 35:1050–1064
Gunderson CA,O’Hara KH,Campion CM,Walker AV,Edwards NT(2010)Thermal plasticity of photosynthesis:the role of acclimation in forest responses to a warming climate.Global Change Biol 16:2272–2286
Hamerlynck E,Knapp AK(1996)Photosynthetic and stomatal responses to high temperature and light in two oaks at the western limit of their range.Tree Physiol 16:557–565
Han FZ,Yang BY,Fan GY,Xia WJ,Ma XD(2015)Identi fi cation of 11 species of bark beetles and their galleries in natural coniferous forests in Qinghai province.For PestDis 34(6):11–16(Abstract in English)
Heroult A,Lin YS,Bourne A,Medlyn BE,Ellsworth DS(2013)Optimal stomatal conductance in relation to photosynthesis in climatically contrastingEucalyptusspecies under drought.Plant Cell Environ 36:262–274
Hikosaka K,Shigeno A(2009)The role of Rubisco and cell walls in the interspeci fi c variation in photosynthetic capacity.Oecologia 160:443–451
Hikosaka K,Ishikawa K,Borjigidai A,Muller O,Onoda Y(2006)Temperature acclimation of photosynthesis: mechanisms involved in the changes in temperature dependence of photosynthetic rate.J Exp Bot 57:291–302
IPCC(2013)Climate change 2013:the physical science basis.Cambridge University Press,Cambridge,p 1535
Kroner Y,Way DA(2016)Carbon fl uxes acclimate more strongly to elevated growth temperatures than elevated CO2concentrations in a northern conifer.Global Change Bio 22:2913–2928
Limousin JM,Bickford CP,Dickman LT,Pangle RE,Hudson PJ,Boutz AL,Gehres N,Osuna JL,Pockman WT,McDowell NG(2013)Regulation and acclimation of leaf gas-exchange in a pi?on-juniper woodland exposed to three different precipitation regimes.Plant Cell Environ 36:1812–1825
Lin YS,Medlyn BE,Ellsworth DS(2012)Temperature responses of leaf net photosynthesis:the role of component processes.Tree Physiol 32:219–231
Luo Y(2007)Terrestrial carbon-cycle feedback to climate warming.Annu Rev Ecol Evol S 38:683–712
Ma F,Zhao CM,Milne RI,Ji MF,Chen LT,Liu JQ(2010)Enhanced drought-tolerance in thehomoploid hybrid speciesPinus densata:implication for its habitat divergence from two progenitors.New Phytol 185:204–216
Mao JF,Wang XR(2011)Distinct niche divergence characterizes the homoploid hybrid speciation ofPinus densataon the Tibetan Plateau.Am Nat 177(4):424–439
Matteo GD,Perini L,Atzori P,Angelis PD,Mei T,Bertini G,Fabbio G,Mugnozza GS(2014)Change in foliar carbon isotope composition and seasonal stomatal conductance reveal adaptive traits in Mediterranean coppices affected by drought.J For Res 25(4):839–845
Niu S,Li Z,Xia J,Han Y,Wu M,Wan S(2008)Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China.Environ Exp Bot 63:91–101
Poorter H,Niinemets U,Poorter L,Wright IJ,Villar R(2009)Causes and consequences of variation in leaf mass per area(LMA):a meta-analysis.New Phytol 182:565–588
Reddy AR,Chaitanya KV,Vivekanandan M(2004)Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants.J Plant Physiol 161:1189–1202
Sendall K,Reich PB,Zhao CM,Hou JH,Wei XR,Stefanski A,Rice K,Rich RL,Montgomery RA(2015)Acclimation of photosynthetic temperature optima of temperate and boreal tree species in response to experimental forest warming.Global Change Bio 21:1342–1357
Smith NG,Dukes JS(2013)Plant respiration and photosynthesis in global-scale models:incorporating acclimation to temperature and CO2.Global Change Biol 19:45–63
Vasseur F,Violle C,Enquist BJ,Granier C,Vile D(2012)A common genetic basis to the origin of the leaf economics spectrum and metabolic scaling allometry.Ecol Lett 15:1149–1157
Wang D,Heckathorn SA,Hamilton W,Frantz J(2014)Effects of CO2on the tolerance of photosynthesis to heat stress can be affected by photosynthetic pathway and nitrogen.Am J Bot 1:34–44
Way DA,Oren R(2010)Differential responses to changes in growth temperature between trees from different functional groups and biomes:a review and synthesis of data.Tree Physiol 30:669–688
Way DA,Sage RF(2008)Thermal acclimation of photosynthesis in black spruce[Picea mariana(Mill.)B.S.P.].Plant Cell Environ 31:1250–1262
Way DA,Yamori W(2014)Thermal acclimation of photosynthesis:on the importance of adjusting our de fi nitions and accounting for thermal acclimation of respiration.Photosynth Res 119:89–100
Yamori W,Noguchi K,Hikosaka K,Terashima I(2009)Coldtolerant crop species have greater temperature homeostasis of leaf respiration and photosynthesis than cold-sensitive Species.Plant Cell Environ 50:203–215
Zhang S,Li Q,Ma K,Cheng L(2001)Temperature-dependent gasexchange and stomatal/non-stomatal limitation to CO2assimilation ofQuercus liaotungensisunder mid-day high irradiance.Photosynthetica 39:383–393
Zhang Q,Chen JW,Li BG,Cao KF(2009)The effect of drought on photosynthesis in two epiphytic and two terrestrial tropical fern species.Photosynthetica 47(1):128–132
Zhang XW,Wang JR,Ji MF,Milne RI,Wang MH,Liu JQ,Shi S,Yang SL,Zhao CM(2015)Higher thermal acclimation potential of respiration but not photosynthesis in two alpinePiceataxa in contrast to two lowland congeners.PLoS ONE 10(4):e0123248
Zhao CM,Chen LT,Ma F,Yao BQ,Liu JQ(2008)Altitudinal differences in the leaf fi tness of juvenile and mature alpine spruce trees(Picea crassifolia).Tree Physiol 28:133–141
Zou XK,Zhai PM,Zhang Q(2005)Variations in droughts over China:1951–2003.Geophys Res Lett 32:L04707.doi:10.1029/2004GL021853
 Journal of Forestry Research2018年2期
Journal of Forestry Research2018年2期
- Journal of Forestry Research的其它文章
- Measurement of lumber moisture content based on PCA and GSSVM
- Characterization of mean stem density, fi bre length and lignin from two Acacia species and their hybrid
- Theoretical modeling of the effects of temperature and moisture content on the acoustic velocity of Pinus resinosa wood
- The properties of fl ax fi ber reinforced wood fl our/high density polyethylene composites
- Risks involved in fecal DNA-based genotyping of microsatellite loci in the Amur tiger Panthera tigris altaica:a pilot study
- Genecological zones and selection criteria for natural forest populations for conservation:the case of Boswellia papyrifera in Ethiopia