
Dead standing pine trees in a boreal forest landscape in the Kalevala National Park,northern Fennoscandia:amount,population characteristics and spatial pattern
2017-10-17 08:31:22TimoKuuluvainenTuomasAakalaandGergelyrkonyi
Forest Ecosystems 2017年3期
Timo Kuuluvainen,Tuomas Aakalaand Gergely Várkonyi
Dead standing pine trees in a boreal forest landscape in the Kalevala National Park,northern Fennoscandia:amount,population characteristics and spatial pattern
Timo Kuuluvainen1*,Tuomas Aakala1and Gergely Várkonyi2
Abstract
Background:After their death,Scots pine trees can remain standing for decades and sometimes up to 200 years,forming long-lasting and ecologically important structures in boreal forest landscapes.Standing dead pines decay very slowly and with time develop into ‘kelo’trees,which are characterized by hard wood with silvery-colored appearance.These kelo trees represent an ecologically important,long lasting and visually striking element of the structure of natural pine-dominated forests in boreal Fennoscandia that is nowadays virtually absent from managed forest landscapes.
Methods:We examined and mapped the amount,structural features,site characteristics and spatial distribution of dead standing pine trees over a ten hectare area in an unmanaged boreal forest landscape in the Kalevala National Park in Russian Viena Karelia.
Results:The mean basal area of dead standing pine trees in the forested part of the landscape was 1.7 m2?ha?1and the estimated volume 12.7 m3?ha?1.From the total number of standing dead pine trees 65%were kelo trees,with a basal area of 1.1 m2?ha?1and volume of 8.0 m3?ha?1,the remainder consisting of standing dead pines along the continuum between a recently dead tree and a kelo tree.Overall,standing dead pines were distributed throughout the study area,but there was a tendency towards spatial clustering up to<100 m distances.Standing dead pines were most commonly situated on flat ground or in the mid slope in the local topography.In addition,standing dead pines contributed to substrate diversity also by commonly having charred wood and broken tops.Based on the presence of dead pine snags in different stage of transition from a recently dead pine to a kelo with silvery surface,it seems evident that the process of kelo recruitment was continuously in action in the studied landscape.
Conclusions:Kelo trees are an omnipresent feature in natural pine-dominated forest landscapes with important contribution to forest structural and substrate diversity.Because of their longevity and extremely slow turnover dynamics and importance for biodiversity,protection of vulnerable kelo tree populations,and ensuring their continuous recruitment,should be of high priority in forest restoration and sustainable management.
Keywords:Boreal forest,Coarse Woody debris,Dead trees,Forest dynamics,Kelo,Pinus sylvestris,Snag,Tree mortality
Background
In boreal Fennoscandia,large dead-standing silvery-colored Scots pine trees are called kelo trees or kelos(Niemel? et al.2002).They are an omnipresent and visually spectacular feature of the structure of pristine pine-dominated forests(Fig.1;Keto-Tokoi and Kuuluvainen 2014).Formation and turnover of kelo trees,as an outcome of natural forest dynamics,is an extremely slow process.In the Fennoscandian boreal forest,it is not uncommon for pines to live for 400–500 years or more(e.g.,Leikola 1969;Zackrisson 1977;Engelmark et al.1994),and following their death,it may take decades for the standing dead pines to develop the characteristic appearance of a kelo tree(Leikola 1969).While standing dead trees in general are well known to be an important habitat structure for a number of species in different groups of organisms,kelos form a further distinct habitat structure for a smaller number of specialists(Niemel? et al.2002;Tikkanen et al.2006;Spribille et al.2008;Vehkaoja et al.2016;Santaniello et al.2017).Importantly,when the kelo trees eventually fall over they become another special type of habitat structure for different species(Niemel? et al.2002).
For a pine to develop into a long-standing kelo tree there seems to be several pre-requisites.First,long-standing dead trees commonly have died gradually,after decadeslong period of slow growth(Niemel? et al.2002;Venugopal et al.2015),likely influenced by senescence,and various disturbances and stressors(Cailleret et al.2017).During this long process that eventually leads to the death of a tree,the wood material impregnates with decay-resisting chemicals(Venugopal et al.2015,2016).Similarly fire scarring of the tree during its lifetime appears to lead to increased impregnation of the wood material at the base of the stem(pers.obs.).This makes the decay of wood very slow and a pine tree can stand upright from some decades to hundreds of years after its death.Over time,the bark and fine branches fall off,and the kelo tree get their characteristic greyish silvery appearance(see Fig.1).What makes standing dead pines stand out from the other common tree species in the European boreal forests is that similar long-term development is not known to occur with the other main tree species.Hence,standing dead pine dynamics are essential for the production of a these specific structural features and habitats.

Fig.1 A typical intact silvery-colored kelo tree,a dead pine tree,in the Kalevala National Park in Russian Viena Karelia.Photograph by Timo Kuuluvainen
The formation,amounts and population dynamics of kelos in natural forests of boreal Fennoscandia are little studied and thus the dynamics of this important habitat feature are poorly understood.Rouvinen et al.(2002)studied the process of trees dying standing in a pine dominated landscape in the area of current Kalevala National Park in Russian Viena Karelia.The 66 kelos,with dated death years,had been standing dead for an average time of 81 years.Two of the oldest kelos aged 257 and 234 years since tree death(Rouvinen et al.2002).Based on this small sample of dated death years,Rouvinen et al.(2002)estimated that the emergence rate of kelos would have been about one new kelo tree per hectare per decade.
In the same Kalevala National Park,Karjalainen and Kuuluvainen(2002)found that the volume of kelo trees(>10 cm dbh)varied from 17.7 to 29.6 m3?ha?1depending on site fertility type.Lampainen et al.(2004)examined a 350-ha pine-dominated forest 31 years after fire,where the mean standing kelo volume was 25.6 m3?ha?1,comprising 26.8%of total dead wood volume.In the study by Karjalainen and Kuuluvainen(2002),pine kelo trees comprised of 30.7%,34.7%and 49.3%of total dead wood pool in mesicVaccinium-Myrtillus-type,dryishEmpetrum-Myrtillustype and dryEmpetrum-Cladina-type sites,respectively(Karjalainen and Kuuluvainen 2002).
Nowadays kelo trees have become rare in boreal forest landscapes wherever humans have been present(Niemel? et al.2002).One reason is that kelo trees are excellent dry firewood.Therefore,they have been through times extensively utilized for all kinds of heating purposes ever since the first hunter-gatherers.While kelo trees have largely disappeared from populated areas already more than 100 years ago(Kalliola 1966),the onset of industrial forest management has led to decline in kelo populations also in more remote parts of Fennoscandia.Because of the shortrotation cycles used in boreal even-aged forest management,there is simply not enough time for kelo trees to emerge in the course of forest development.
Kelo trees,and the standing dead pines that precede their formation,are important for biodiversity(Niemel? et al.2002).However,the natural properties,dynamics and spatial structure of kelo trees and their populations have been scarcely studied,especially at larger spatial scales,as former studies typically used relatively small sample plots(e.g.Karjalainen and Kuuluvainen 2002).Accordingly,the purpose of this study was to examine the amount,structure and spatial distribution of kelo trees in a boreal forest landscape in the Kalevala National Park in Russian Viena Karelia.In addition,we examined whether the occurrence of kelo trees can be explained by environmental factors such as local topography and tree density,and amount of stumps from past selective logging;the area has been the source of construction wood for a small nearby village that has been inhabited since the sixteenth century.Even if not in a pristine state,we believe the current forest structure to be a result of a typical forest use in boreal Fennoscandia in large,sparsely populated areas,prior to population growth and the onset of industrial forest management.To our knowledge this is so far the first landscape-level spatially explicit mapping and examination of kelo trees in an unmanaged forest landscape in northern Fennoscandia.
Methods
Study area
The study was carried out in the roadless Vienansalo wilderness area in the Kalevala National Park in Russian Viena Karelia.The study area is located in the eastern part of the Fennoscandian shield in the middle boreal vegetation zone(Fig.2;Kalela 1961;Ahtiet al.1968).The study area was located in the north-western side of Lake Venehj?rvi(Russian name is Sudnozero;65°00′N,30°05′E).The terrain is characterized by small hills and the average elevation of the study site is 155 m a.s.l.The growing season is approximately 140 days,and effective temperature sum 900 d.d.(+5°C threshold).Annual precipitation is 650 mm,of which half falls as snow.The underlying parent rock is mostly composed of gneiss with a high proportion of biotite.The study area is characterized by a thin layer of moraine between parent rock and soil(Atlas Karelskoy ASSR 1989;Gromtsev 1998;Gorkovets et al.2000).
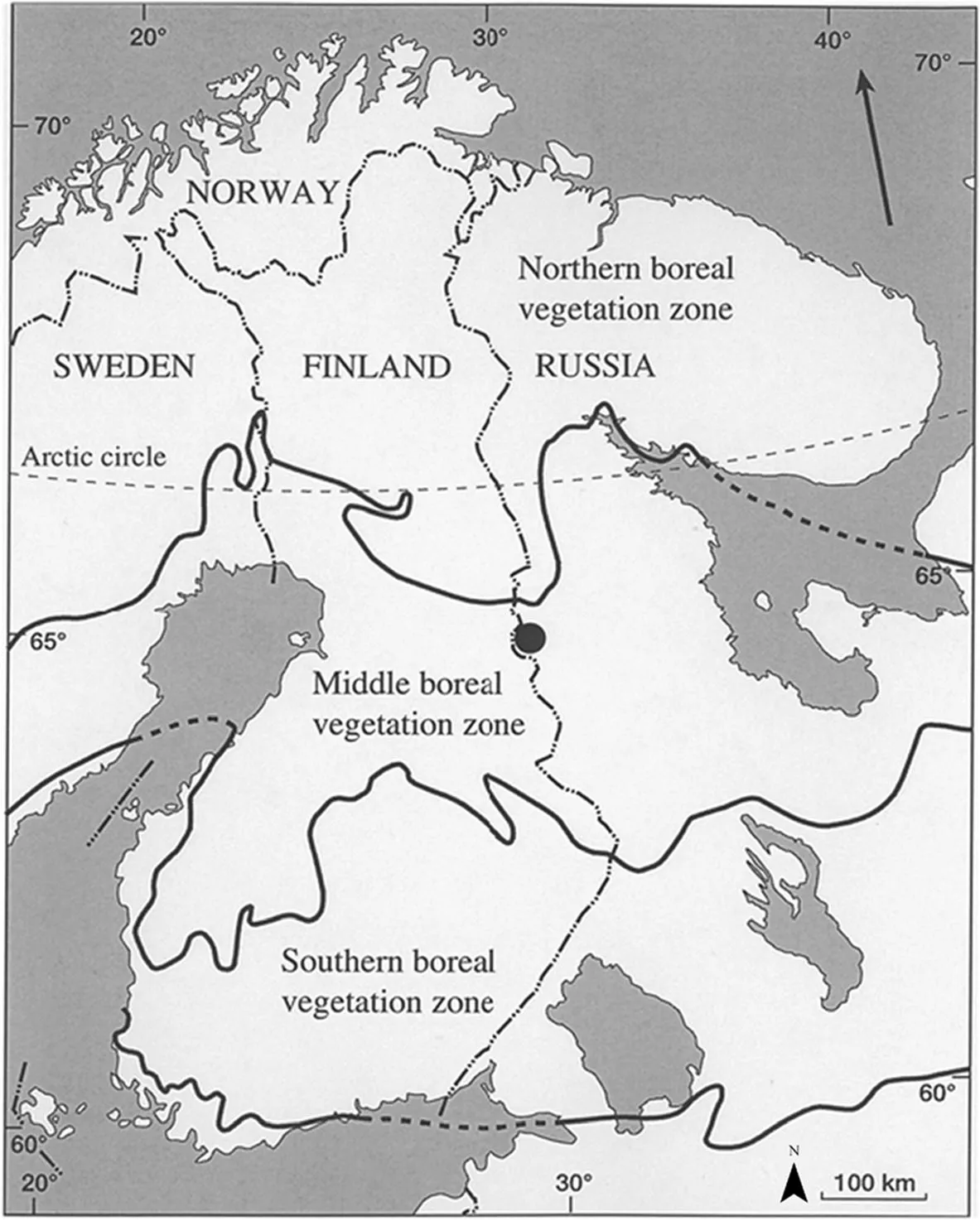
Fig.2 The geographic location of the study area in the Kalevala National Park in Russian Viena Karelia
Anthropogenic influence has been low in the area,especially during the past 80 years.Human activity was probably most pronounced in the late 19th and early twentieth century.The population of the nearby Venehj?rvi village probably peaked in the early 1940s,when there were around 300 inhabitants in the village,which is located approx.5 km distance from the study area.The use of natural resources was subsistence based.Trees were felled mostly for local building and heating purposes.Accordingly signs of human activity such as cut stumps can be seen in some areas as signs of past selective loggings.Working partly in the same study area,Karjalainen and Kuuluvainen(2002)recorded on average 14 cut stumps per hectare(range 0–45 ha?1).TheVenehj?rvi village was abandoned during the soviet era and currently only one family lives there year round.The Kalevala National Park was founded in 2006 and it covers an area of 740 km2.
Sampling and measurements
We selected the study area in the forested landscape near Lake Venehlampi by first locating the northeastern corner of the study area from satellite imagery.The exact location was determined so that the plot was fully located within the forest-mire mosaic,away from the direct human influence that is often most prominent around water bodies,but within a reasonable walking distance from Lake Venehlampi.Although subjectively located on the satellite image,the exact location of the study area was determined without prior examination in the field.
We conducted the field sampling in May–June 2015.In the field,we first delineated the study plot as follows:we laid out an approximately 25 m×25 m grid over a rectangular area of 200 m×500 m(ca.10 ha).The location of the grid nodes was determined,using a consumer-grade handheld GPS device.At each node,we measured the basal area of live and dead trees with a relascope,separately for each tree species,viz.pine,spruce and deciduous trees.We marked this grid in the field,and inventoried stumps from old selective loggings,by recording their numbers within each grid cell.Due to the inaccuracies of uncorrected GPS locations,the grid node locations contained considerable uncertainty.However,the purpose at this point was to avoid subjective selection of the grid points,rather than to delineate a perfect grid.
Within the study plot,we then mapped all standing dead pine stems exceeding 1.3 m in height,and with a diameter over 10 cm at 1.3 m height(DBH).We mapped standing dead pines,using a Trimble GeoXM GPS receiver that facilitates postprocessing of the recorded locations.Simultaneously,we also recorded the accurate locations of the grid nodes for postprocessing.Due to technical issues with the VRS-correction data(provided by Geotrim Ltd.,Vantaa,Finland)we were able to correct the location of 257 out of the total of 355 dead standing pine trees recorded.The location accuracy was 2.9 m(i.e.,66%of the time the location accuracy is at least this good),which we consider sufficient for our analyses.Grid node locations were all corrected,and their location accuracy was 1.9 m.
For each mapped standing dead pine,we recorded its DBH,and height,as well as the following characteristics:percentage of bark remaining(1%–100%),presence of charred wood(0/1),fire scar(0/1),cavity(0/1)and silvery color of the trunk(0/1),and whether the tree was snapped or intact(0/1).Each of these trees were classified into one of five decay classes,based on their visual appearance,mainly the presence or absence of needles,and branches of different order(Aakala 2010).In addition,the basal area of the forest surrounding each recorded tree was measured with a relascope separately for pine,spruce and deciduous trees.Kelo trees were defined as standing dead pine trees which had grey or silvery stem surface.Most of these trees had only little bark remaining;for example,80%of kelo trees had less than 20%bark remaining.
Data computations
While we use the basal area and number of standing dead pines in our present analyses,we also computed kelo volumes for comparisons with other studies.For volume computations,we used volume integrals of taper equations,developed for pine in Finland(Laasasenaho 1982).The advantage of using taper equations(in addition to taking stem taper into account)is that the volume can easily be computed for trees of any height.However,this requires information on height of the intact tree,which together with tree diameter is then used to determine the stem taper.To obtain intact heights for snapped dead pines,we fitted N?slund’s(1936)height curve to an independent DBH-height dataset from the same region,from Kuuluvainen et al.(2002),and used this to predict the height of the intact trees(i.e.,snapped trees prior to snapping).Volume integrals were then computed,using functions for the R statistical computing environment(R Core Team 2016),by Aakala(2017).
For assessing the influence of local stand condition,past selective logging,and small-scale topographic position,we created a set of predictor variables by interpolating data recorded at the grid nodes,supplemented with elevation data at each tree.Using corrected location data from kelo tree measurements and the grid nodes,we created a digital elevation model for the study area,using the 3DAnalyst extension in ArcGis 10.3(spherical variogram model,using elevation data from 12 nearest points as the search radius).From this elevation model,we created a raster oftopographic positions,using the topographic position index in the Topography toolbox(with a rectangular neighborhood of 50 m;Dilts 2015),into valley,toe slope,flat,mid slope,upper slope and ridge.We used ordinary kriging(spherical semivariogram model with 12 points neighborhood)to create a raster of basal area values,and a raster of selective logging intensity(parameters as in basal area raster,but with 6 nearest point neighborhood),from the number of cut stumps.
For the spatial analyses,we divided the study area into hexagonal cells with 10 m as the length of the side of the hexagon(and area of 260 m2)(Fig.3).The size of the hexagons was chosen as a compromise between the loss of spatial resolution,and the number of hexagon cells with zero kelo trees.After removing the hexagon cells that fell within the open mire in the study area,we had 387 grid cells.For each cell,we computed the number of kelos,and their basal area,and extracted the elevation,topographic position,stand basal area(live trees,and live+dead trees separately),and the number of stumps as the mean value in the cell,or as the most frequent value in the case of the topographic position.
Analyses
We used the basal area measurements at the grid node points to calculate the average basal area and its distribution(absolute and proportional)into living and dead trees as well as by tree species in the study area.The relation between occurrence of kelo trees and elevation in topography,density of living trees and number of old stumps from selective logging were analyzed at the hexagonal grid cell scale(Fig.3).Because the data proved to be autocorrelated,we used spatial autoregressive modeling(SAR)to examine which of the environmental variables were related to kelo tree occurrence(basal area).We used Moran’sIstatistics to summarize the spatial autocorrelation pattern in kelo tree abundance.The SAR modelling and spatial analyses were carried out with the SAM software(Rangel et al.2010).
Results
The overall mean basal area was 12.3 m2?ha?1,of which 88.4%was made up by living trees and 11.6%by dead standing trees(Table 1).Pine made up the majority of living tree volume(58.7%)followed by spruce(17.1%),while living deciduous trees was a minor component(1.4%)in terms of basal area in the landscape.
Altogether,we mapped 354 standing dead trees larger than 10 cm at dbh in the study area.Of these 319(90.1%)were pine trees,13(3.7%)were aspen,12(3.4%)Norway spruce and 10(2.8%)goat willow.The mean density of standing dead pines was 35.8 per hectare,excluding the small treeless bog area(Fig.3).The estimated mean basal area and volume of standing dead pines was 1.7 and 12.7 m3per hectare,respectively;excluding the small treeless bog area.The size distribution of these dead trees was fairly normal,and trees between 20 and 30 cm at dbh were most abundant(Fig.4).The mean dbh was 25.8 cm and the maximum 56 cm.
A total of 208 of the dead pine trees(65.2%)were recorded as having the characteristic silvery appearance,and were classified as kelo trees.Fire scars and charred wood were found on 130 of the dead pines(40.8%of trees),171 trees(48.0%)were broken or snapped,and 166(52%)dead pines were snapped.Cavities were recorded in 13(3.8%)of the standing dead pines.Kelo trees with low or no bark cover were the most abundant,but also standing dead trees with most of the bark remaining were quite common(Fig.5).
The standing dead pines were most often located on flat ground or on mid slope in the local topography(Fig.6).The distribution of kelos in the local topography was asfollows:valley 3.3%,toe slope 6.5%,flat 41.3%,mid slope 34.2%,upper slope 8.2%and ridge 2.2%.
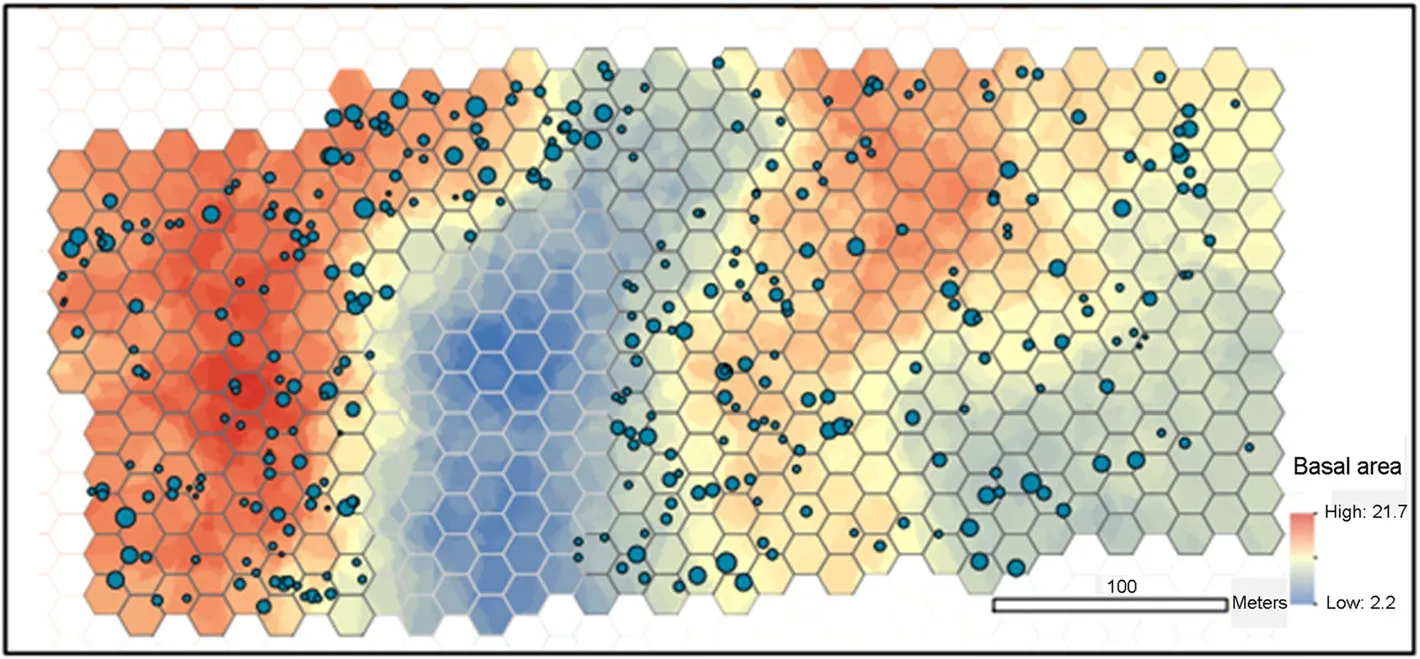
Fig.3 The study area showing the spatial pattern of the standing dead pine trees.Size of the filled circles is proportional to the size the dead standing pine trees.The background color show the spatial distribution of total tree basal area(including both live and dead trees)along with the hexagonal grid cells used in interpolations.Cells on the open mire were omitted and are shown with light grey borders

Table 1 Basal areas and tree species proportions by tree species and divided into living and dead tree components on the study are based on the systematic sample of relascope measurements
At least some standing dead pines were found throughout the study area,except for the near-open peat bog with only some small stunted pine trees(Fig.6).However,the density of standing dead pines varied considerably across the study area and some notable clustering was visually apparent in parts of the study area(Fig.6).This was further confirmed in a spatial autocorrelation analysis,indicating significant clustering of standing dead pine basal area at distances less than 100 m(Fig.7).
The impact of past human activity was described as the number of old selectively logged tree stumps,the number of which was computed for the 260 hexagonal grid cells.There were,on average,13.5 cutting stumps per hectare in the study area.46%of the grid cells(and studied area)had no cutting stumps and 91%had≤1 stump.The spatial distribution of stumps was very uneven and concentrated in two parts of the study area(data not shown).
The spatial autoregressive(SAR)modeling,using the gridded data,indicated that the standing dead pine basal area was best explained by the combined basal area of living and dead trees(probably reflecting the overall longterm local competition environment at the site,see Fig.3)and elevation in the local topography,while the number of cut stumps was not a significant predictor of basal area standing dead pines(Table 2).
Discussion
The mean volume of standing dead pines in our study area was 12.7 m3?ha?1.This is comparable but somewhat lower than the values obtained in the study of Karjalainen and Kuuluvainen(2002),also carried out in the Kalevala National Park,with the volume of kelo trees varying from 17.7 to 29.6 m3?ha?1the highest value found in the poorestEmpetrum-Cladina-type.Also working in the area that is currently part of the Kalevala National park,Lampainen et al.(2004)examined a 350-ha pine dominated forest 31 years after fire,where mean standing kelo tree volume was 25.6 m3?ha?1.It is evident that in unmanaged northern forests,kelo trees can comprise a significant share of total dead tree volume;for example,in the study of Karjalainen and Kuuluvainen(2002)this was 30%–50%depending on site type(Karjalainen and Kuuluvainen 2002;Table 1 and Fig.3).The occurrence of standing dead pine basal area increased with the combined basal area of living and dead trees and decreased with elevation in the local topography(Table 2).The former relationship may reflect the overall long-term local competition environment at the site,whilethis and the(rather weak)elevation effect may be related to more favorable regeneration and growth conditions at lower compared with and high drier locations in the local topography(Fig.3).The fact that the number cut stumps was not a significant predictor of basal area standing dead pines can be due to low number of cut trees and long time elapsed from cuttings.Earlier studies in the same region have shown that stands selectively cut in the past may not significantly differ in their structure from more natural stands(Lilja and Kuuluvainen 2005).

Fig.4 Diameter distribution of the dead standing pine trees in the study area

Fig.5 Distribution of the dead standing pine trees by bark cover percentage classes
Standing dead pines,with their abundance,longevity,structural diversity and slow population dynamics,are an essential feature and long-lasting structural component of boreal forest landscapes in boreal Fennoscandia(Fig.8).Because of slow decay,they obviously strongly affect and stabilize carbon dynamics of the ecosystem,and to lesser effect the nutrient dynamics(as the stemwood is fairly nutrient-poor;Helmisaari and Siltala 1989).They also significantly contribute to the structural and substrate diversity of the forest.Therefore,it is not surprising that many organisms use standing dead pines as habitats and are even specialized to utilize this structure and its substrate.For example,a large number of specialized epiphytic crustosecalicioidlichens(also known asCaliciales)and some fungi live on the surface of kelo trees(Niemel? et al.2002;Vehkaoja et al.2016).

Fig.6 The spatial distribution of the dead standing pine trees together with the variation in topography in the study area.The local topography variation range was 115–132 m a.s.l.Cells on the open mire were omitted and are shown with light grey borders
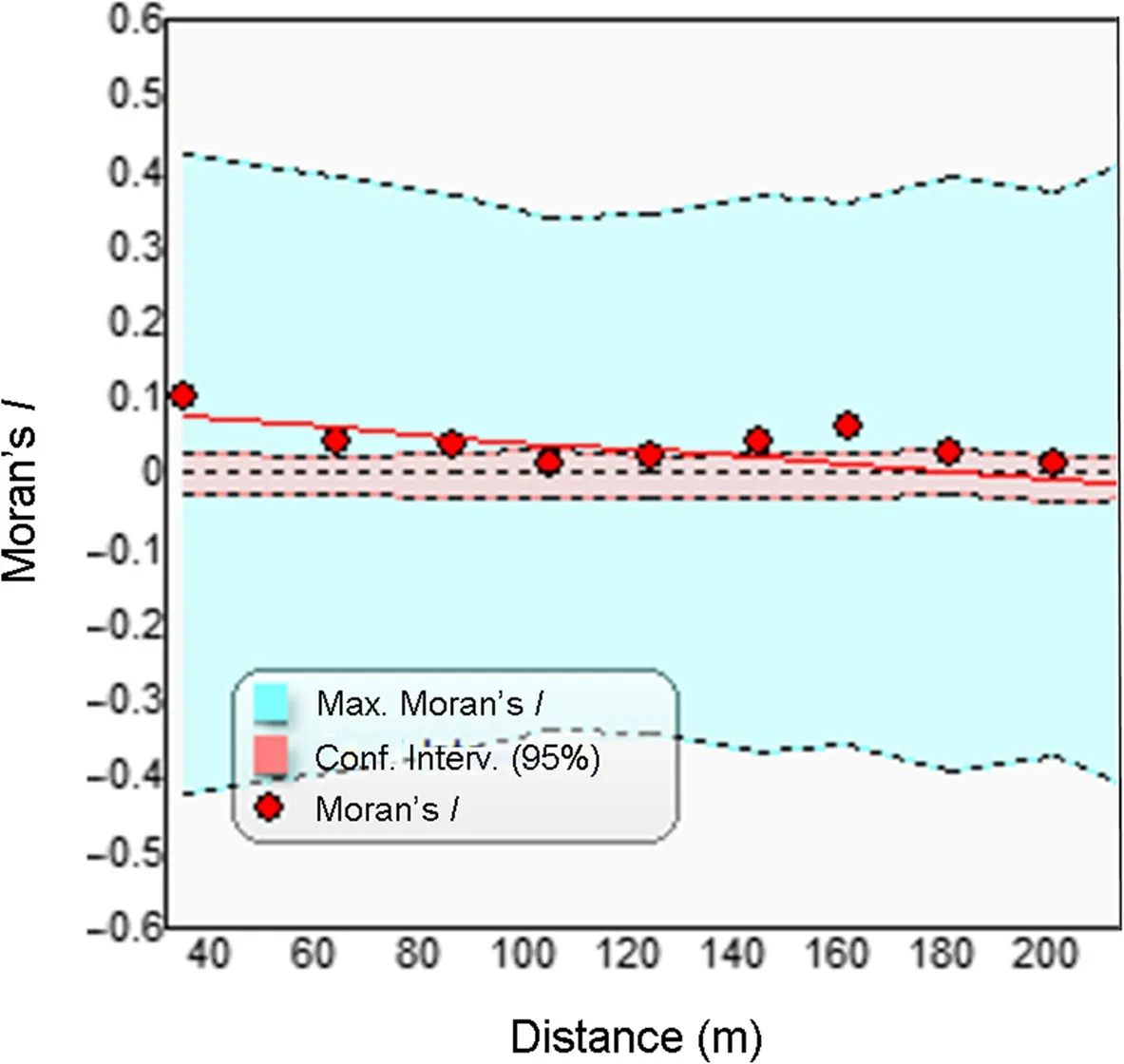
Fig.7 Spatial autocorrelation analysis(Moran’s I)of dead pine basal area
In forest fires,standing dead pines and kelo trees easily catch fire and burn partly and therefore they may comprise a significant amount of charred wood,to which some lichen species are specialized(L?hmus and Kruustuk 2010;Fig.8).In our study area 41%of the kelos had been charred reflecting past fires in the landscape(Lehtonen and Kolstr?m 2000).Substrate type diversity is also increased by disturbances such as broken tree tops,which were recorded for 52%of the kelos.Also woodpeckers,for example the big Black Woodpecker(Dryocopus martius),are known to make their nesting cavities in kelo trees(Fayt 2003),thus providing habitat for a number of other cavity nesting birds and animals,including Tengmalm’s Owl(Aegolius funereus),Goldeneye(Bucephala clangula),Swift(Apus apus),bats and the Siberian Flying Squirrel(Pteromys volans).Thus,kelo trees can also potentially be an important structural component contributing to the“cavity web”in these forests(cf.Martin and Eadie 1999;Martin et al.2004;Cooke and Hannon 2011).However,in our data only 12(3.8%)kelos were reported to have a cavity.It may be that kelo wood is so hard that only trees with decayed heartwood are attractive for woodpecker nesting.Large hollow and broken kelos can also be used for nesting by large owls such as the Great Grey Owl(Strix nebulosa)(Fig.8).

Table 2 Results of the spatial autoregressive model(SAR)for the hexagonal grid-wise basal area(BA)of dead standing kelo trees(n=387,F=5.056,p=0.002)
The dispersal ecology and population dynamics of the specialized kelo species are likely adapted to the spatiotemporal population dynamics of the kelo resource they live on.In our study area,kelo trees were distributed throughout the area(excluding the treeless bog area),but there was also some spatial clustering.This result is in accordance with that of Rouvinen et al.(2002),looking at tree mortality dynamics in the same area but on a larger pine-dominated forest landscape.We did not measure the time since tree death for the kelos,but based on the decay class distribution that is known to be linked to time since tree death(Aakala 2010),it is clear that kelos are continuously formed in the studied landscape.This,together with the results of Rouvinen et al.(2002),indicate that the kelo formation process is in action across the larger forest landscape in the Kalevala National Park.
The continuous formation and tendency to spatial clustering suggest that kelo occurrence and dynamics are characterized by some kind of spatial-temporal continuum,meaningthat kelo habitats are continuously available over relatively short distances to their accompanying species such as epiphytic lichens(Boch et al.2013).An interesting further question would be how this pattern is reflected in the dispersal abilities of kelo-associated species and how these species might be dependent of patterns of kelo availability in time and space.

Fig.8 Examples of different kelo structures contributing to forest structural and substrate diversity.a A fairly recently died partly debarked tree(often fall of bark is accelerated by woodpecker foraging).b Old kelo tree with fire scarring and charred wood in the trunk base.c A kelo with a cavity.The tree was made hollow by the Boreal Carpenter Ant(Camponotus herculeanus).d A large hollow broken kelo tree;such hollow broken trees are used for nesting by e.g.the Great Grey Owl.Photographs by Timo Kuuluvainen
Conservation of kelo trees and their dynamics pose a particular challenge,especially in regard to their reintroduction to forests.In even-aged forest management short rotation cycles used and there is simply not enough time for kelo trees to emerge in the course of forest development.This is because of the slow formation of kelos:it can be roughly estimated that after a clearcut it can take 500 to 1000 years before a kelo tree population and its dynamics are restored.Therefore,leaving some retention pine trees on clearcuts can be recommend as one possible way to enhance kelo tree recruitment in managed forests(Gustafsson et al.2012;Santaniello et al.2016).However,this may not be sufficient,as the formation of long-standing kelo trees might be partially related to the occurrence of surface fires,which were historically common but now practically absent from pine dominated landscapes in Fennoscandia(Wallenius 2011).Surface fire probably leads to changes in the chemical composition of the base of the tree stem and hence increased decay resistance and longevity of kelo trees.In the absence of forest fires,kelo population dynamics might be experiencing changes also in the conservation areas.Thus,restoration burnings in protected areas might be beneficial for restoration of kelo tree populations and their dynamics.Managing for kelo trees and conserving their specialized species can be regarded as an ultimate conservation and restoration challenge in boreal forest landscapes.
Acknowledgements
This work was carried out in the framework of the EBOR-project funded by the Academy of Finland(proj.no.276255).We also thank the Finnish Environment Institute for helping in financing this research and Mr.Santeri Lesonen from the Venehj?rvi village for hospitality and help in practical arrangements.
Authors’contributions
TK participated in planning,field work and data analyses of the study.He was mainly responsible of the writing of the manuscript.TA participated in planning,field work,data analyses and writing of the manuscript.GV participated in the planning and organization of the research and commented the manuscript.All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Author details
1Department of F2orest Sciences,University of Helsinki,P.O.Box 27,FI-00014 Helsinki,Finland.Finnish Environment Institute,Natural Environment Centre,Friendship Park Research Centre,Lentiirantie 342B,FI-88900 Kuhmo,Finland.
Aakala T(2010)Tree mortality and deadwood dynamics in late-successional boreal forests.Dissertationes Forestales 100:41 http://www.metla.fi/dissertationes/df100.htm.Accessed 10 Mar 2017
Aakala T(2017)R functions for computing tree stem volumes.Figshare.https://doi.org/10.6084/m9.figshare.4616479.v1.Accessed 10 Mar 2017
AhtiT,H?met-AhtiL,Jalas J(1968)Vegetation zones and their sections in northwestern Europe.Ann Bot Fenn 5(3):169–211
Atlas Karelskoy ASSR(1989)Glavnoe upravlenie geodezii i kartografii pri Sovete Ministrov SSSR.Moscow,p 40(in Russian)
Boch S,PratiD,Hessenm?ller D,Schulze ED,Fischer M(2013)Richness of lichen species,especially of threatened ones,is promoted by management methods furthering stand continuity.PLoS One 8(1):e55461.doi:10.1371/journal.pone.0055461
Cailleret M,Jansen S,Robert EMR,Desoto L,Aakala T,Antos JA,Beikircher B,Bigler C,Bugmann H,Caccianiga M,?ada V,Camarero JJ,Cherubini P,Cochard H,Coyea MR,?ufar K,Das AJ,Davi H,Delzon S,Dorman M,Gea-Izquierdo G,Gillner S,Haavik LJ,Hartmann H,Here? A-M,Hultine KR,Janda P,Kane JM,Kharuk VI,Kitzberger T,Klein T,Kramer K,Lens F,Levanic T,Linares Calderon JC,Lloret F,Lobo-Do-Vale R,Lombardi F,López Rodríguez R,M?kinen H,Mayr S,Mészáros I,Metsaranta JM,Minunno F,Oberhuber W,Papadopoulos A,Peltoniemi M,Petritan AM,Rohner B,Sangüesa-Barreda G,Sarris D,Smith JM,Stan AB,Sterck F,Stojanovi? DB,Suarez ML,Svoboda M,TognettiR,Torres-Ruiz JM,Trotsiuk V,Villalba R,Vodde F,Westwood AR,Wyckoff PH,Zafirov N,Martínez-Vilalta J(2017)A synthesis of radial growth patterns preceding tree mortality.Glob Change Biol 23:1675–1690
Cooke HA,Hannon SJ(2011)Do aggregated harvests with structural retention conserve the cavity web of old upland forests in the boreal plains?For Ecol Manag 261(3):662–674.doi:10.1016/j.foreco.2010.11.023
Dilts TE(2015)Topography tools for ArcGIS 10.1.University of Nevada Reno.http://www.arcgis.com/home/item.html?id=b13b3b40fa3c43d4a23a1a0 9c5fe96b9.Accessed 10 Mar 2017
Engelmark O,Kullman L,Bergeron Y(1994)Fire and age structure of Scots pine and Norway spruce in northern Sweden during the past 700 years.New Phytol 126(1):163–168
Fayt P(2003)Population ecology of the three-toed woodpecker under varying food supplies.University of Joensuu,PhD Dissertations in Biology,Joensuu,No.21
Gorkovets VY,Rayevskaya MB,Lukashov AD(2000)The geology of the proposed Kalevala,Tuulij?rvi,Koitajoki and Tolvaj?rvi national parks.In:Heikkil? R,Heikkil? H,Polevoi A,Yakovlev E(eds.).Biodiversity of old-growth forests and its conservation in northwestern Russia.North Ostrobothnia Regional Environment Centre.Regional Environmental Publications 158,p 159–172
Gromtsev AN(1998)Inventory of natural complexes and ecological feasibility study of Kalevala National Park.Preprint of the paper presented at the session of the Research Board of the Forest Research Institute,Karelia Research Centre,RAS,27 November 1997.Forest Research Institute,Karelian Research Centre,Russian Academy of Sciences,Petrozavodsk,p 56
Gustafsson E,Baker SC,Bauhus J,Beese WJ,Brodie A,Kouki J,Lindenmayer DB,Lohmus A,Martinez Pastur G,Messier C,Neyland M,Palik B,Sverdrup-Thygeson A,Volney VJA,Wayne A,Franklin JF(2012)Retention forestry to maintain multifunctional forests:a world perspective.Bioscience 62(7):633–645
Helmisaari H-S,Siltala T(1989)Variation in nutrient concentrations of Pinus sylvestris stems.Scand J For Res 4:443–451
Kalela A(1961)Waldvegetationszonen Finnlands und ihre klimatische Paralleltypen.Arch Soc Vanamo 16(Suppl.):65–83
Kalliola R(1966)The reduction of the area of forests in natural condition in Finland in the light of some maps based upon national forest inventories.Ann Bot Fenn 3(3):442–448
Karjalainen L,Kuuluvainen T(2002)Amount and diversity of coarse woody debris within a boreal forest landscape dominated by Pinus sylvestris in Vienansalo wilderness,eastern Fennoscandia.Silva Fenn 36(1):147–167
Keto-Tokoi P,Kuuluvainen T(2014)Primeval forests of Finland.Cultural history,ecology and conservation.Maahenki,Porvoo,p 301
Kuuluvainen T,M?ki J,Karjalainen L,Lehtonen H(2002)Tree age distributions in old-growth forest sites in Vienansalo wilderness,eastern Fennoscandia.Silva Fenn 36(1):169–184
Laasasenaho J(1982)Taper curve and volume functions for pine,spruce and birch.Commun Inst For Fenn 108:1–74
Lampainen J,Kuuluvainen T,Wallenius TH,Karjalainen L,Vanha-Majamaa I(2004)Long-term forest structure and regeneration after wildfire in Russian Karelia.J Veg Sci 15(2):245–256
Lehtonen H,Kolstr?m T(2000)Forest fire history in Viena Karelia,Russia.Scand J For Sci 15:585–590
Leikola M(1969)Havaintoja m?nnyn paksuuskasvun loppumisesta ja puiden keloutumisesta Inarin Lapissa.Silva Fenn 3(1):50–61(in Finnish)
Lilja S,Kuuluvainen T(2005)Structure of old Pinus sylvestris dominated forest stands along a geographic and human impact gradient in mid-boreal Fennoscandia.Silva Fenn 39(3):407–428
L?hmus P,Kruustuk K(2010)Lichens on burnt wood in Estonia:a preliminary assessment.Folia Cryptogam Est 47:37–41
Martin K,Eadie JM(1999)Nest webs:a community-wide approach to the management and conservation of cavity-nesting forest birds.For Ecol Manag 115(2–3):243–257
Martin K,Aitken KEH,Wiebe KL(2004)Nest sites and nest webs for cavity-nesting communities in interior British Columbia,Canada:nest characteristics and niche partitioning.Condor 106(1):5–19
N?slund M(1936)Skogsf?rs?ksanstaltens gallringsf?rs?k i tallskog.Meddelanden fr?n Statens Skogsf?rs?ksanstalt 29:169
Niemel? T,Wallenius T,Kotiranta H(2002)The kelo tree,a vanishing substrate of specified wood-inhabiting fungi.Pol Bot J 47(2):91–101
R Core Team(2016)R:a language and environment for statistical computing.R Foundation for Statistical Computing,Vienna,Austria
Rangel TF,Diniz-Filho JAF,Bini LM(2010)SAM:a comprehensive application for spatial analysis in Macroecology.Ecography 33(1):46–50
Rouvinen S,Kuuluvainen T,Siitonen J(2002)Tree mortality in a Pinus sylvestris dominated forest landscape in Vienansalo wilderness,eastern Fennoscandia.Silva Fenn 36(1):127–145
Santaniello F,Djupstr?m L,Ranius T,Rudolphi J,Widenfalk O,Weslien J(2016)Effects of partial cutting on logging productivity,economic returns and dead wood in boreal pine forest.For Ecol Manag 365:152–158
Santaniello F,Djupstr?m LB,Ranius T,Weslien J,Rudolphi J,Thor G(2017)Large proportion of wood dependent lichens in boreal pine forest are confined to old hard wood.Biodivers Conserv 26(6):1295–1310
Spribille T,Thor G,Bunnell FL,Goward T,Bj?rck CR(2008)Lichens on dead wood:species-substrate relationships in the epiphytic lichen floras of the Pacific northwest and Fennoscandia.Ecography 31(6):741–750
Tikkanen OP,Martikainen P,Hyv?rinen E,Junninen K,Kouki J(2006)Red-listed boreal forest species of Finland:associations with forest structure,tree species,and decaying wood.Ann Zool Fenn 43(4):373–383
Vehkaoja M,Nummi P,Rikkinen J(2016)Beavers promote calicioid diversity in boreal forest landscapes.Biodivers Conserv 26:579–591
Venugopal P,Julkunen-Tiitto R,Junninen K,Kouki J(2015)Phenolic compounds in Scots pine heartwood:are kelo trees a unique woody substrate?Can J For Res 46(2):225–233
Venugopal P,Junninen K,Linnakoski R,Edman M,Kouki J(2016)Climate and wood quality have decayer-specific effects on fungal wood decomposition.For Ecol Manag 360:341–351
Wallenius T(2011)Major decline in fires in coniferous forests–reconstructing the phenomenon and seeking for the cause.Silva Fenn 45(1):139–155
Zackrisson O(1977)Influence of forest fires on the north Swedish boreal forest.Oikos 29(1):22–32
*Correspondence:Timo.Kuuluvainen@helsinki.fi
1Department of Forest Sciences,University of Helsinki,P.O.Box 27,FI-00014 Helsinki,Finland
Full list of author information is available at the end of the article
?The Author(s).2017 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0
International License(http://creativecommons.org/licenses/by/4.0/),which permits unrestricted use,distribution,and
reproduction in any medium,provided you give appropriate credit to the original author(s)and the source,provide a link to the Creative Commons license,and indicate if changes were made.
Received:23 March 2017 Accepted:29 June 2017

- Forest Ecosystems的其它文章
- Development and evaluation of robust tree biomass equations for rubber tree(Hevea brasiliensis)plantations in India
- Climate change effects in the Western Himalayan ecosystems of India:evidence and strategies
- Small mammal community response to early meadow–forest succession
- Stand dynamics of the drought-affected floodplain forests of Araguaia River,Brazilian Amazon
- Tree species diversity and utilities in a contracting lowland hillside rainforest fragment in Central Vietnam
- Conspecific negative density dependence in American beech