
Small mammal community response to early meadow–forest succession
2017-10-17 08:31:21LinasBaliauskasAuraepukienandLaimaBaliauskien
Forest Ecosystems 2017年3期
Linas Bal?iauskas,Au?ra ?epukien? and Laima Bal?iauskien?
Small mammal community response to early meadow–forest succession
Linas Bal?iauskas*,Au?ra ?epukien? and Laima Bal?iauskien?
Abstract
Background:With farmland afforestation becoming common policy in many European Union countries,we studied how early forest succession(from meadow to young stand)influences small mammal species composition,diversity,abundance and biomass.Despite numerous investigations into forest succession,almost no attention has been given to the small mammal community change in the early-successional forest ecosystems,starting with the pre-forest habitat and ending with stand formation and the establishment of tree dominance.We compared small mammal communities in meadows at the initial stage of regrowth(with saplings less than 10 cm in height),in young forest(5–10 years old)and more advanced forest(15–20 years)in both cases of human-induced forest succession,where the trees had been planted,and natural forest succession,where natural regrowth of meadows had occurred.
Results:The greatest diversity of small mammal species was recorded in the meadow(H=2.95),with a lower diversity found in the young forest(H=2.61)and even lower in the advanced forest(H=2.04),the last habitat being the most monodominantic.The order of species dominance fromMicrotussp.(M.arvalis,M.agrestis),Myodes glareolus,Apodemus flavicollis,Sorex araneus,A.agrariusin the meadow changed toM.glareolus,S.araneus,M.arvalis,M.agrestisin the young forest and toM.glareolus,A.flavicollis,S.araneusin the advanced forest.The lowest relative abundance of small mammals was recorded in the meadow(18.19±2.27 ind.Per 100 trap-days),withMicrotusvoles being the most abundant.Relative abundance was higher in the young forest(22.72±2.25 ind.Per 100 trap-days),withMyodes glareolusbeing the most abundant(7.59±0.96 ind.Per 100 trap-days)and at its highest in the advanced forest(23.91±2.77 ind.Per 100 trap-days),again withM.glareolusbeing the most abundant(15.54±2.35 ind.Per 100 trap-days).
Conclusions:Thus,our analysis suggests that that during early meadow-forest succession,the diversity of the small mammal community declines–the number of species decreases as typical meadow species are lost due to the transformation of the habitat and one or a few species became dominants.However,the relative abundance of the small mammals increases.Biological indices of small mammal communities differed between natural and humaninduced meadow-forest succession.
Keywords:Afforestation,Voles,Mice,Diversity,Abundance,Biomass
Background
Farmland afforestation has recently become a common policy in European Union countries,involving not only subsidies,but also research into its contribution to carbon sequestration,land uptake and impact on local biodiversity(Kotecky 2015).In the early succession stage,when the forest is still not yet dominated by tree canopies,there is a high production yield by various plant components and the habitat is characterised by high complexity and wide food webs(Swanson et al.2010).Comparing planted forests to natural growth forests,there is a clear trade off between the produced goods and ecosystem services and a decrease of biological diversity(Carnus et al.2006).Yet still,the area of planted forest continues to increase by ca.2%annually and evaluation of their significance to preservation of biological diversity is not clear(Brockerhoff et al.2008).
Human Induced Succession(hereafter HS)occurs throughout the post-Soviet countries,with seedlings planted in abandoned fields and meadows as part of a policy of afforestation.In Lithuania,the actions of theForest Cover Enlargement Programme have accelerated in recent years in particular.For example,forest cover in the territory of Lithuania had increased by 104,000 ha during the previous decade(Butkus et al.2013),compared to 44,900 ha in the decade before(Kavaliauskien? and Tarvydien? 2005).Simultaneous in 1992–2002 the area of arable land fell by 118,000 ha and that of meadows and natural pastures by 89,100 ha.
Forests may also develop in localities of former meadows as a process of Natural Succession(hereafter NS)when abandoned arable land and/or hay meadows are recolonized by shrubs and eventually become forests.This has become common in Lithuania and other Baltic countries since 1990,where following land reform,less intensive farming has led to a decrease in agricultural areas(Aleknavi?ius and Aleknavi?ius 2010)and 4000–5000 ha of abandoned land has seen natural forest regrowth on an annual basis(Lithuanian Forest Cover Enlargement Programme 2002).Currently,forest and newly afforested land occupies 33.3%of the territory of the country(Butkus et al.2013).
In both cases(HS and NS),the early succession stages are characterised as being diverse in species,processes and structure(Swanson et al.2010).It has been shown that sampling in all habitats is important for understanding small mammal community changes in forest–farmland ecosystems(Panzacchi et al.2010).For example,quite unexpectedly,newly afforested sites may harbour a poor small mammal diversity in comparison to habitat undergoing deforestation–for the first 15 years at least,newly afforested habitats in China were found to be dominated by agricultural pest species(Raoul et al.2008).For the first four years of afforestation,small mammals do not tend to react to it,as could be the case in other types of disturbances that fragment their habitat(Johnson et al.2002).
Despite small mammals being recognized as biological indicators of sustainable forest management in the boreal zone(Pearce and Venier 2005),most research has focussed on changes in their communities during postdisturbance forest succession:after fires,clear cutting and logging(i.e.,Gashwiler 1970;Kirkland 1990;Sullivan et al.1999;Briani et al.2004;Swanson et al.2010;Urban and Swihart 2011;Borchert et al.2014).Only a few investigations have dealt with changes in the small mammal community during meadow-to-forest succession stages(Atkeson and Johnson 1979;Huntly and Inouye 1987;Swihart and Slade 1990).In Lithuania,meadowto-forest succession and its impact on small mammals has also received relatively little investigation(but see Bal?iauskas and Angelstam 1993;Jasiulionis et al.2011;?epukien? and Jasiulionis 2012).
The aim of this study was to understand how early forest succession(from meadow to young stand)influences small mammal species composition,diversity,abundance and biomass.Previous investigations into forest succession have not focussed on how the community of small mammals changes in the early-successional forest ecosystems,starting with the pre-forest habitat and ending with stand formation and the establishment of tree dominance.Thus,we tested if changes are the same under HS(Human Induced Sucession,where forests have been planted)and earlyNS (NaturalSuccession,where natural regrowth of unused meadows has occurred).Thus we assessed if afforestation programs(which revert unused land into forest plantations)negatively affect small mammal communities.
Hypothesis H1 was that early forest succession diminishes small mammal species diversity.Hypothesis H2 was that small mammal biomass under early forest succession is nevertheless maintained,as a loss of species diversity is compensated by a higher abundance,and hence biomass,of the dominants.Hypothesis H3 was that both types of early forest succession(HS and NS)have the same influence on the small mammal community.
Methods
Study area
Investigations into small mammal community changes during early forest succession stages were carried out in temperate mixed forests(Lithuania)in June–September of 2007–2008 and 2010–2012,with additional data also in September 2013.Small mammal species in Lithuania have no clearly expressed cyclic fluctuations of abundance(Bal?iauskas and Angelstam 1993;Bal?iauskas and Ju?kaitis 1997;Bal?iauskas 2005).For both natural forest regrowth and planted forest,investigations were conducted in three types of habitat that can be regarded as covering early forest succession:1)meadow in the initial stage of regrowth,with trees less than 20 cm in height or shrubs(hereafter referred to as ‘meadow’),2)former meadows now covered by trees of approximately 5–10 years old with a canopy still open(hereafter referred to as‘young forest’,and 3)former meadow now covered by developing birch-spruce forest,trees approximately 15–20 year old and with a closed canopy(hereafter referred to as‘advanced forest’).Additional information about the habitats is presented in Additional file 1(Tables S1 and S2).The localities for the studies of the three habitat types,for both the HS and NS,were in close proximity to each other(Fig.1).
Small mammal trapping methods
In June–August of 2007–2008 and 2010–2012,small mammals were trapped in each habitat by live traps set in three lines with 25 traps per line,each trap five metres apart.Live-traps were left in place for three days and the traps checked twice per day.The live-trapped animals were marked,weighed,described and then released,dataused to determine species composition and abundance.In September of 2007–2008 and 2010–2013,small mammals were trapped by a standard linear snap-trap method(Bal?iauskas 2004)to determine species composition,abundance,age structure and breeding data by dissecting trapped animals.Again,for each habitat,traps were set in three lines,25 per line and each trap five metres apart.The snap-traps were left in position for three days,and the traps were checked every day.Both trapping methods were used to determine the community’s species composition,diversity,dominance and relative abundance.

Fig.1 Location of study sites:a–Human Induced Sucession,b–Natural Succession,yellow colour denotes meadows,blue colour young forest,and green colour advanced forest.HS:meadow 1.26 ha,55°58′35.19″N,23°48′11.17″E;young forest 2.8 ha,55°59′13.57″N,23°47′56.78″E;advanced forest 6 ha,55°59′11.5″N,23°48′1.27″E.NS:meadow 1.3 ha,55°44′39.35″N,25°45′7.03″E;young forest 2 ha,55°44′42.34″N,25°45′19.15″E;advanced forest 1.6 ha,55°44′51.56″N,25°45′20.93″E
The abundance of all small mammals trapped was assessed using a relative index,i.e.the number of individuals trapped per 100 traps in the first day(ind.Per 100 trap-days),where “day”means 24-h period from morning to next morning.The trapping effort in the habitats of HS was 3890 trap?days?1,while in the habitats of NS it was 2159 trap?days?1.Biomass(g?ha?1)was expressed as the sum of the body mass of all individuals trapped in a line of 25 snap traps from the same habitat;such a line corresponds to 1 ha,as stated in Kleemola and S?derman(1993).
In both live-and snap-trapping,line placement was chosen randomly in the first year,fitting all traps of the straight line to the same habitat,and the same placement used for all investigation period.
Sample size
A total of 1591 small mammals belonging to 11 species of the orders Insectivora and Rodentia were trapped between 2007 and 2013(1044 individuals of 11 species in HS,and 547 individuals of 10 species in NS habitats).Without respect to succession type(HS or NS),all 11 small mammal species were recorded in the meadow and ten species each in the young forest and the advanced forest.More detailed sample size information for the habitats of HS and NS is presented in Table 1.
Statistical analyses
The species composition of small mammal communities was determined using Shannon’s diversity indexHon log2base and Simpson’s species dominance indexc(Brower and Zar 1984).The indices were calculated using StatEcol software(Ludwig and Reinolds 1988).The significance of small mammal diversity differences was estimated using the DivOrd program 1.90 version.H±SD calculations were done in the DOSBox ver.0.74 environment(Tóthmérész 1993).The Rényi diversity index(Tóthmérész 1998)was used to test if the small mammal diversity differences were significant with respect to habitat and time.
To compare small mammal communities,the family of diversity indices is represented graphically using Rényi diversity profiles,where the values of parameter α are from 0 to 4.When α =0,the Rényi diversity index is equal to the logarithm of the number of species;when α =1,the Rényi index is equal to Shannon’sH;when α = 2,the Rényi index reflects Simpson’s dominanceindex;when α =3 and 4,the Rényi profiles show higher degree diversity indices(Tóthmérész 1998;Carranza et al.2007).

Table 1 Sample size(number of registered species and number of trapped small mammal individuals)in the habitats under human-induced and natural early meadow-forest succession
The effect of the habitat,year,season and succession type(HS or NS)on small mammal community parameterswasassessed using multidimensionalstatistics methods(factorial ANOVA),and pair-wise differences were tested using Student’s t-tests by comparing more than two sets;the Bonferoni correction was used(Zar 1999;StatSoft 2013).Significance of theMyodestoMicrotusratio in meadow,young forest and advanced forest was tested using chi-square statistics.Calculations were done with Statistica for Windows(StatSoft 2013).
Results
The following species of small mammals were registered in the habitats of early forest succession:common shrew(Sorex araneus),pygmy shrew(S.minutus),water shrew(Neomys fodiens),yellow-necked mouse(Apodemus flavicollis),striped field mouse(A.agrarius),house mouse(Mus musculus),harvest mouse(Micromys minutus),bank vole(Myodes(Clethrionomys)glareolus),common vole(Microtus arvalis),root vole(M.oeconomus)and field vole(M.agrestis).
Effect of early forest succession on small mammal species composition and diversity
Several significant differences were found in small mammal species composition when comparing the habitats of early forest succession.The meadow habitat was characterised by the highest small mammal diversity.There was no clear dominance–the number of trapped individuals is similar in the six most abundant species(Table 2).M.arvaliswere trapped most frequently,whileM.glareolus,S.araneus,andA.flavicolliswere less numerous.M.musculuswas characteristic to this stage only.The number ofM.oeconomustrapped in the meadow was higher than in other habitats,but its overall proportion in the small mammal community was not great.
In the young forest,the dominance ofM.glareoluswas already clear,with individuals of this species accounting for one third of all individuals trapped.However,the proportions of the other small mammal species that were abundant in the meadows still remained high in the young forest.
The advanced forest was strongly dominated byM.glareolus,which accounted for more than half of all small mammals trapped.A.flavicollisandS.araneuswere also numerous,but the proportions of other small mammal species had decreased and did not exceed 5%.
The order of species dominance changed with succession:fromMicrotusvoles(M.arvalis,M.agrestis),Myodes glareolus,Apodemus flavicollis,Sorex araneus,A.agrariusin the meadow,toM.glareolus,S.araneus,M.arvalis,M.agrestisin the young forest,and toM.glareolus,A.flavicollis,S.araneusin the advanced forest.Dynamic ofMyodes/Microtusratio is shown in the Fig.2,and supports hypothesis H1 about the species change.Ratio change to theMyodesbehalf along with initial forest succession is significant(χ2=1265.4,df=2,P<0.0001).

Table 2 Species composition of small mammals in early meadow-forest succession(N:number of individuals,%:species share in the habitat)
Thus,hypothesis H1 was confirmed(i.e.small mammal diversity under meadow-forest succession diminishes due to the growing dominance ofM.glareolusand decreasing abundance ofM.arvalisand other meadow dwellers).
Effect of early forest succession on small mammal abundance and biomass
For both types of succession combined,the average small mammal abundance was(mean±SE)18.19±2.27(0–40) ind.Per 100 trap-days in the meadow,22.72 ± 2.25(0–40)ind.Per 100 trap-days in the young forest and 23.91 ± 2.77(4–56)ind.Per 100 trap-days in the advanced forest(Table 3).Long-term abundance differences between habitats were not significant(meadow–young forest,t=1.41,meadow–advanced forest,t=1.60,young forest–advanced forestt=0.33).The average abundance of infrequent species did not significantly differ between habitats(Table 3).The average abundance ofM.glareolusin the meadow was lower than in the young or advanced forest,and average abundance in the young forest was lower than in the advanced forest.The average abundance ofMicrotusvoles,on the contrary,was at its lowest in the advanced forest,i.e.lower than in either the young forest or meadow;their abundance in the meadow and in the young forest did not differ.Thus,the recorded changes in the average abundances of small mammals between habitats could largely be attributed to the changes in the most abundant species.
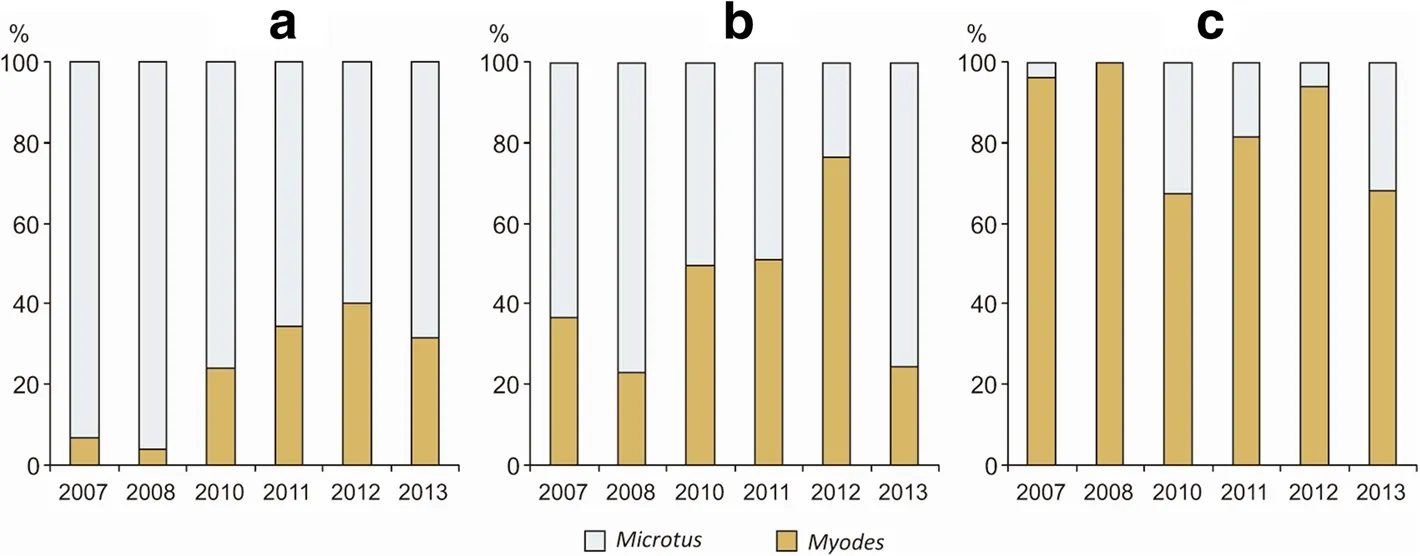
Fig.2 Change of the Myodes to Microtus sp.ratio in meadows(a),young forests(b)and advanced forests(c)in 2007–2013
Irrespective of the type of succession,the average biomass of small mammals was 399.0 ± 68.6 g?ha?1in the meadow,424.1 ± 83.1 g?ha?1in the young forest and 367.9 ± 50.9 g?ha?1in the advanced forest,with the differences between habitats not significant(meadow–young forest,t=0.23,meadow–advanced forest,t=0.58,young forest–advanced forestt=0.36).
The biomass ofM.glareoluswas significantly lower in the meadow than in the young forest(t=2.06,P<0.05)and advanced forest(t=3.97,P<0.001).The biomass ofM.arvalisin the meadow and young forest did not differ significantly,but it was lower in the advanced forest(t=2.61,P=0.012 compared with the meadow,andt=2.17,P<0.05 compared with the young forest).Similar differences were observed in the biomass of allMicrotusvoles,with the lowest value being in the advanced forest(t=2.94,P=0.012 compared with the meadow,andt=2.17,P<0.05 compared with the young forest),and no difference in biomass between the meadow and the young forest.
Thus,hypothesis H2wasconfirmed (i.e.despite changes in the small mammal species composition,biomass did not decrease in the later successional stages,particularly due to the higher numbers,hence biomass,ofM.glareolus).
The total effect of “succession type”(Human Induced Succession or Natural Succession)or “habitat”, “year”and “species”on the biomass of small mammals was significant(ANOVA,r2=0.62,F161,240=2.46,P<0.0001).Though the factor “year”was not significant it exhibited a trend(F=2.02,P=0.13).Furthermore,the effect of the interactions “succession type”× “year”was significant(F=5.12,P<0.01),as was the interaction among“year”× “species”(F=2.79,P< 0.005)and the threeway interaction in succession among “type”× “year”× “-species”(F=2.32,P=0.01).

Table 3 Relative abundance of small mammal species in early meadow-forest succession,irrespective of succession type(X–mean relative abundance,individuals per 100 trap-days;SE–standard error;Min–max–minimum and maximum values)
The biomasses of various small mammal species were affected by a different number of factors.An analysis of the total effect of “habitat”,“succession type”,“year”and“season”revealed that for changes in the biomass ofA.agrarius(ANOVAr2=0.58,F12,54=6.21,P<0.0001)only the trapping-month was significant (F= 13.01,P<0.0001).The biomasses of the other three abundant small mammal species were affected by succession type,year and month:S.araneus(r2=0.58,F12,54=9.74,P<0.0001;F=39.54,F=10.99,F=14.38,respectively,allP<0.0001),M.glareolus(r2=0.562,F12,54=7.31,P<0.0001;F=6.46,P=0.013,F=3.25,P=0.012 andF=15.02,P<0.0001,respectively),A.flavicollis(r2=0.51,F12,54=4.61,P<0.0001;F=8.12,P<0.01,F=6.54,P<0.0001 andF=3.46,P=0.014,respectively).
The biomass ofMicrotusvoles(ANOVAr2=0.39,F12,54=2.79,P<0.005)was significantly affected by habitat(F=3.61,P<0.05)and trapping-year,i.e.cyclicality(F=3.11,P=0.015).The biomass ofM.arvalis(r2=0.52,F12,54=4.79,P<0.0001)was due to succession type(F=7.21,P<0.01),habitat(F=4.90,P=0.011),and year(F=4.16,P=0.003),while the biomass ofM.agrestis(r2=0.42,F12,54=3.28,P=0.0013)was due to year(F=3.10,P=0.016)and month(F=4.12,P=0.005),but not habitat(F=1.59,P=0.21).
Small mammal diversity under natural and human-induced early meadow-forest succession
In general,small mammal diversity was at its highest in the meadow,lower in the young forest and at its lowest in the advanced forest(Fig.3).In the case of HS,the differences in the small mammal species diversity between these habitats were significant(meadow–young forest,t=4.55,meadow–advanced forest,t=12.21,young forest–advanced forestt=8.65,allP<0.0001;Fig.3a).In the case of NS,small mammal diversity in the young forest was significantly higher than in the meadow,but small mammal diversity in the advanced forest did not differ from that in the meadow or in the young forest(Fig.3b).
In the HS meadow,S.araneus(24.2%of all individuals trapped)andA.flavicollis(19.9%)were the dominant species.Microtusvoles,namelyM.arvalis(12.5%),M.agrestis(11.3%),andM.oeconomus(9.8%),were subdominants.In the NS meadow,dominant species wereM.arvalis(26.1%)andM.glareolus(25.7%),subdominants wereA.agrariusandM.agrestis,constituting 11.3%and 13.6%respectively.
In the HS young forest,M.glareolus(36.8%)was the dominant species.S.araneus,M.agrestis,andM.arvalis(17.2%,12.4%and 10.7%respectively)were subdominants.In the NS young forest,the proportion of dominant species was 26.2%forM.glareolusand 22.7%forM.arvalis.Subdominants wereM.agrestisandA.agrarius(13.4%and 12.8%respectively).
M.glareoluswas the dominant species in both HS(61.9%)and NS(40.7%)advanced forests.Subdominants in HS advanced forests wereA.flavicollisandS.araneus(15.5%and 12.2%),withS.araneusandM.arvalisthe subdominants in NS advanced forest(with 12.7%and 13.6%respectively).
In the habitats undergoing HS,the lowest species diversity was in the advanced forest(Shannon’sH=0.95–2.09;averageH=1.73)and the highest was in the meadow(H=2.92).In this respect,the young forest(H=2.56)was closer to the meadow than to the advanced forest.In the habitats undergoing NS,the highest small mammal diversity was recorded in the young forest(H=2.24).Small mammal diversity was more variable in habitats undergoing HS(H=1.73–2.92)than NS(H=2.54–2.61).Large differences were observed between forest stands under HS and NS,natural succession preserving more diverse small mammal community(H=1.73 andH=2.61 respectively).

Fig.3 Rényi diversity in habitats undergoing early meadow-to-forest succession.Small mammal diversity profiles in the habitats of human-induced succession are presented in a,natural succession in b,and the averaged data of both succession types in c.One small mammal community can be considered more variable than the other if the Rényi diversity profiles do not intersect
In the habitats undergoing HS,the small mammal community was monodominantic in the plantedadvanced forest(Simpson’sc=0.43)and polydominantic in the meadow(c=0.15).Under NS,the small mammal community was polydominantic in all three habitats:advanced forest(c=0.24),meadow and young forest(c=0.20).Thus,the small mammal dominance indices differed considerably depending on the succession type(natural or induced),thereby allowing the rejection of hypothesis H3.
Differences in small mammal abundance and biomass depending on the meadow-forest succession type(NS or HS)
The abundance of small mammals in meadows was mostly dependent onMicrotusvoles.Other abundant species wereM.glareolusin the NS meadow andA.flavicollisin the HS meadow.The average small mammal abundance was significantly higher in meadow undergoing NS(Table 4).
The average abundance of small mammals in the young forest undergoing HS and NS did not differ,and the relative abundances of the most abundant species–M.glareolus,Microtusvoles andS.araneus– did not differ either(Table 4).The only significant difference was the higher abundance ofM.arvalisin the NS young forest.
The average small mammal abundance in the advanced forest undergoing HS was almost three times the abundance in the advanced forest under NS.The difference was due to a greater abundance ofM.glareolusandA.flavicollis,which were over compensating the decrease in abundance ofMicrotusvoles(Table 4).
Changes in small mammal biomass during the early meadow-to-forest succession mostly depended on the type of succession.In case of HS,the biomass ofM.glareolusincreased significantly from 19.5 g?ha?1in the meadow to 160.5 g?ha?1in the planted young forest(t29=3.05,P< 0.005)and to 258.6 g?ha?1in the planted advanced forest(t=6.54,P=0.0001 compared with the meadow,andt=2.75,P=0.01 compared with the young forest).The biomass of allMicrotusvoles did not differ between the meadow and young forest(t=0.87,P=0.39),but it was significantly lower in the advanced forest(t=2.06,P<0.05 compared with the meadow,andt=2.81,P<0.01 compared with the young forest).The biomass ofM.agrestiswas 32.9 g?ha?1in planted advanced forest,whileM.arvaliswas not trapped at all in the planted advanced forest.
Though changes in the biomass of small mammals during NS were less significant,the total biomass of small mammals did decrease significantly in the natural advanced forest as compared to the meadow(t12=2.26,P<0.05),this being due to the decrease in the biomass ofMicrotusvoles from 331.6 g?ha?1in the meadow to 68.6 g?ha?1in the advanced forest(t=2.59,P=0.023).
Thus,we can further reject hypothesis H3:comparing induced and natural early meadow-forest succession,we found differences in species composition,diversity,relative abundance and biomass.
Discussion
Afforestation of unused land is not the only solution for its restoring habitats–extensive grazing and clearing of scrubland is proposed by some scientists(Lasanta et al.2015).Despite this knowledge,the main land changes inEurope are still those related to cropland/grassland processes and afforestation;deforestation should be mentioned as a historical perspective(Fuchs et al.2015).Planted forest in Europe covered 32 million hectares in 2001,equalling 17%of the world’s forest plantations.Forest ecosystems may be diverse in the early succession stage,developed after disturbance or after replacing of the initial advanced forest(Swanson et al.2010).In the case of forestdevelopment by planting in former meadows or agricultural land,woodland development can be as short as 15 years,whereas natural succession in abandoned fields could result in shrublands,not forest stands,even after more than 50 years(Huntly and Inouye 1987).

Table 4 Relative abundance of small mammal species in the meadow,young forest and advanced forest,depending on the succession type(X:mean relative abundance,individuals per 100 trap-days,SE:standard error;significance of HS-NS difference:*:P<0.05,**:P<0.01,***:P<0.001)
It has already been shown that plantation forests can be suitable as habitat,even to some rare and threatened species,mainly birds,amphibians and insects(Brockerhoff et al.2008).The main changes that occur after afforestation relate to vegetation cover(Decocq et al.2005).Planted forest stands are usually inhabited by a lower number of native species compared to native forests,but this number is in most cases greater than in degraded ecosystems.Thus,afforestation of abandoned land(meadows,pastures or agricultural fields)may maintain biodiversity by providing forest habitat,increasing ecotone area and connectivity between habitats(Brockerhoff et al.2008).Small mammal abundance mostly depends on the heterogeneity of habitat and factors such as forest floor,presence of stones,vertical shelter and soil moisture(Carey and Harrington 2001).In the early stage of forest succession,species diversity is high due to the presence of survivor,habitat specialist and opportunist species.However,this phase may be of limited length in planted forests(Swanson et al.2010).
Despite numerous investigations into forest succession,almost no attention has been given to the small mammal community change in the early-successional forest ecosystems,starting with the pre-forest habitat and ending with stand formation and the establishment of tree dominance.According to Fox(1995),a shift in the community structure occurs when the dominating species,after a change of habitat,decrease in number and are replaced by species with better adaptability.Investigation of old field(2–57 years)succession proved that meadow succession leads to small mammal community changes:abundance was low and not dependent on the time of succession,but species diversity depended on plant yield and thus the limiting factor was nitrogen content in the soil(Huntly and Inouye 1987).In Norway,abandoned meadows boasted the highest abundance and diversity of small mammals.In younger meadows,the dominating species wasMicrotus agrestis,while at later stages,the shrubby meadows were dominated byM.glareolus(Panzacchi et al.2010).
In the early successional stages,species richness depends on the presence of tall vegetation and the structural heterogeneity of the forest.Late successional traits have little positive influence,thus a high species richness of small mammals is characteristic to the youngest of forests(Kirkland 1990;Sullivan et al.2000;Ecke et al.2002).Other authors state that species richness of small mammals increases as forests mature to 15–20 years,then decreases thereafter with a minimum achieved at a stand ageofabout40 years(Schoonmakerand McKee 1988;Torre and Diaz 2004).However,migration between the three successional stages-meadow,shrubbymeadow and youngforest-doesoccur(Swihart and Slade 1990).
Changes in the small mammal community after forest fires or disturbance(clearcutting)are well-documented(see Zwolak 2009).In both cases,the diversity of small mammals is similar and the growth in their abundance corresponds to the stand age with a maximum in the mature forest(Fisher and Wilkinson 2005).Mature forest monoculture also supports a low abundance of small mammals in Norway(Panzacchi et al.2010).A maximum diversity of small mammal species is registered at 15 years after a fire(Briani et al.2004),then it declines and reaches a minimum in the 40 year-age forest stands(Schoonmaker and McKee 1988).Dense tree planting shortens the duration of the early succession stage and can reduce species richness(Swanson et al.2010).
Only a few investigations into the response of small mammals to land abandonment and the re-growth of shrubs and trees had been conducted in Lithuania prior to our study.Long term investigations(1981–1990)in meadows with planted spruce seedlings showed that the small mammal community lost a number of species under meadow succession.In one territory undergoing this succession,five to seven small mammal species were trapped in 1981–1983,three to five species in 1984–1985 and only 2–3 species after 1986.As the black alder and birch canopy developed,forest dwelling species replaced meadow species and diversity fell to a minimum.The formerly most abundantM.arvalis(22.7%–79.6%of all trapped small mammals in 1981–1985)lost its position andM.glareolusstarted to dominate(50%–85.7%in 1986–1990)(Bal?iauskas and Angelstam 1993).Abandonment of agricultural land also resulted in dominant species change fromM.arvalisorA.flavicollisin the field fragments toM.glareolusin the re-growing forest patches.M.glareoluswas the only species which successfully adapted to landscape matrix changes after forest regrowth in the abandoned land(?inkūnas and Bal?iauskas 2006).
The results of the current study are not fully comparable with the data from the earlier non-systematic small mammal trapping carried out in similar habitats inLithuania and results are also not consistent.In the earlier study in the meadows experiencing re-growth,three to five species of small mammals were trapped and diversity varied(H= 1.16–2.25),with the dominant species accounting for 30%–66.7%of all trapped individuals.Dominants wereA.agrarius,S.minutusand,in heavy shrubbed meadow,M.glareolus(Bal?iauskas and Ju?kaitis 1997).Relative abundance also varied,ranging from 12 to 44 individuals per 100 trap-nights.
Also,the species diversity of small mammals in the meadows prior to the re-growth was much higher,with 8–13 species trapped(Bal?iauskas and Angelstam 1993;Bal?iauskas and Alejūnas 2011).By contrast,the current study yielded 11 small mammal species trapped in the HS meadow and eight species in the NS meadow.
In previous studies,the number of small mammal species registered in young forest growths was three to nine,with the dominant species beingM.glareolus,A.flavicollisandS.araneus(Ma?eikyt? 2002;Alejūnas and Stirk? 2010;Bal?iauskas and Alejūnas 2011).Results of our study are in good accordance with these published data.In advanced forest in other territories in Lithuania,four to nine small mammal species were registered,withM.glareolusdominating in all cases(Ma?eikyt? 2002;Alejūnas and Stirk? 2010;Bal?iauskas and Alejūnas 2011).Again,the results of our study are similar,with eight small mammal species registered in the advanced forest undergoing HS and 10 species in the advanced forest under NS.
Like many long-term studies,our study has only temporal replication so one must be a bit cautious in generalizing our findings.However,in light of the continuing trend for afforestation in the EU,we feel that it is important that additional studies on how afforestation affects the richness and diversity of small mammal communities be conducted,and that they have both temporal and site replication.
Conclusions
Our results show that the negative effects of early forest succession on small mammal communities are milder in the case of Natural Succession (NS)compared to Human induced Succession(HS).Other long term studies also confirm that the diversity of small mammals is higher in natural stands than in planted forests(Saitoh and Nakatsu 1997).However,a previous study also showed that when the succession started from planting spruce into meadows(HS),the abundance of small mammals did not drop,just the dominant species changed(?inkūnas and Bal?iauskas 2006).More studies like ours are needed to understand how afforestation in the EU may affect the diversity of small mammal populations.
Additional file
Additional file 1:Table S1.Composition of vegetation in meadow(M),young(YF)and advanced(AF)forest in sites of Human Induced(HS)and Natural(NS)succession.Notation according Braun-Blanquet:+?plant sparse,cover up to 1%of area;1-plants cover up to 5%;2-up to 25%;3–25–50%;4–50–75%area.Table S2.Projection of various cover types,cover damages and usage in meadow(M),young(YF)and advanced(AF)forest in sites of Human Induced(HS)and Natural(NS)succession.HS site had slightly better coverage by shrubs and young trees(up to 6 m height).Tree density is higher in the HS site.After thinning,some stumps were left in the HS site.However,grass coverage is similar in both,HS and NS sites.(DOCX 39 kb)
Abbreviations
HS:human-induced forest succession;NS:natural forest succession
Author’s information
LB1 is leading scientist at the Nature Research Centre,head of the Laboratory of Mammalian Ecology,LB2 is senior scientist at the same laboratory.A? obtained PhD in Ecology and Environmental Sciences in 2014,now working as HR manager in business.
Ethics approval
All applicable institutional and/or national guidelines for the care and use of animals were followed.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
LB1 drafted the manuscript and made statistical analysis.A? performed all trappings.LB1,LB2 and A? did all laboratory work(measuring and dissecting small mammals).LB2 commented all manuscript versions.Final manuscript was read and approved by all authors.

Alejūnas P,Stirk? V(2010)Small mammals in northern Lithuania:species diversity and abundance.Ekologija 56:110–115.doi:10.2478/v10055-010-0016-6
Atkeson TD,Johnson AS(1979)Succession of small mammals on pine plantations in the Georgia Piedmont.Am Midl Nat 101:385–392.doi:10.2307/2424604
Aleknavi?ius A,Aleknavi?ius P(2010)Perspectives of Farming Lands Area Preservation in Lithuania.L?ūU mokslo darbai 86:28–36(in Lithuanian)
Bal?iauskas L(2004)Methods of Investigation of Terrestrial Ecosystems.Part I.Animal Surveys,VUL,Vilnius(in Lithuanian)
Bal?iauskas L(2005)Results of the long-term monitoring of small mammal communities in the Ignalina Nuclear Power Plant Region(Drūk?iai LTER site).Acta Zool Litu 15:79–84.doi:10.1080/13921657.2005.10512378
Bal?iauskas L,Angelstam P(1993)Ecological diversity:to manage it or to restore?Acta Ornithologica Lituanica 7:3–15
Bal?iauskas L,Ju?kaitis R(1997)Diversity of small mammal communities in Lithuania(1.A review).Acta Zool Litu Biodiversity 7:29–45.doi:10.1080/13921657.1997.10541423
Bal?iauskas L,Alejūnas P(2011)Small mammal species diversity and abundance in ?agar? Regional Park.Acta Zool Litu 21:163–172.doi:10.2478/v10043-011-0017-z
Borchert MI,Farr DP,Rimbenieks-Negrete MA,Pawlowski MN(2014)Responses of Small Mammals to Wildfire in a Mixed Conifer Forest in the San Bernardino Mountains,California.Bull South Calif Acad Sci 113:81–95.doi:10.3160/0038-3872-113.2.81
Briani DC,Palma ART,Vieira EM(2004)Post-fire succession of small mammals in the Cerrado of central Brazil.Biodivers Conserv 13:1023–1037
Brockerhoff EG,Jactel H,Parrotta JA,Quine CP,Sayer J(2008)Plantation forests and biodiversity:oxymoron or opportunity?Biodivers Conserv 17:925–951.doi:10.1007/s10531-008-9380-x
Brower JE,Zar JH(1984)Field and laboratory methods for general ecology,second edn.wm.c.brown company publishers,Dubuque
Butkus A,Eigirdas M,Kulie?is A,Mik?nait? E,Vi?lenskas D(2013)Lithuanian Statistical Yearbook of Forestry 2013,Lutut?,Kaunas(in Lithuanian)
Carey AB,Harrington CA(2001)Small mammals in young forests:implications for management for sustainability.Forest Ecol Manag 154:289–309.doi:10.1016/s0378-1127(00)00638-1
Carnus JM,Parrotta J,Brockerhoff E,Arbez M,Jactel H,Kremer A,Lamb D,O’Hara K,Walters B(2006)Planted forests and biodiversity.J Forest 104:65–77
Carranza ML,Acosta A,Ricotta C(2007)Analyzing landscape diversity in time:the use of Rényi’s generalized entropy function.Ecol Indic 7:505–510.doi:10.1016/j.ecolind.2006.05.005
?epukien? A,Jasiulionis M(2012)Small mammal community changes during forest succession(Pakruojis district,NE Lithuania).Zool Ecol 22:144–149.doi:10.1080/21658005.2012.739866
Decocq G,Aubert M,Dupont F,Bardat J,Wattez-Franger A,Saguez R,De Foucault B,Alard D,Delelis-Dusollier A(2005)Silviculture-driven vegetation change in a European temperate deciduous forest.Ann For Sci 62:313–323.doi:10.1051/forest:2005026
Ecke F,L?fgren O,S?rlin D(2002)Population dynamics of small mammals in relation to forest age and structural habitat factors in northern Sweden.J Appl Ecol 39:781–792.doi:10.1046/j.1365-2664.2002.00759.x
Fisher JT,Wilkinson L(2005)The response of mammals to forest fire and timber harvest in North American boreal forest.Mammal Rev 35:51–81.doi:10.1111/j.1365-2907.2005.00053.x
Fox BJ(1995)Long-term Studies of Small Mammal Communities from Disturbed Habitats in Eastern Australia.Academic Press,Orlando
Fuchs R,Herold M,Verburg PH,Clevers JG,Eberle J(2015)Gross changes in reconstructions of historic land cover/use for Europe between 1900 and 2010.Glob Chang Biol 21:299–313.doi:10.1111/gcb.12714
Gashwiler JS(1970)Plant and mammal changes on a clearcut in West-Central Oregon.Ecology 51:1018–1026.doi:10.2307/1933628
Huntly N,Inouye RS(1987)Small mammal populations of an old–field chronosequence:successional patterns and associations with vegetation.J Mammal 68:429–435 http://dx.doi.org/10.2307/1381550
Jasiulionis M,?epukien? A,Bal?iauskas L(2011)Small mammal community changes during succession of the planted forest.Acta Zool Litu 22:293–300.doi:10.2478/v10043-011-0035-x
Johnson R,Ferguson JWH,Van Jaarsveld AS,Bronner GN,Chimimba CT(2002)Delayed responses of small-mammal assemblages subject to afforestationinduced grassland fragmentation.J Mammal 83:290–300 http://dx.doi.org/10.1093/jmammal/83.1.290
Kavaliauskien? B,Tarvydien? ME(2005)Changes of agricultural land and forest areas in Lithuania.L?ūU mokslo darbai 67:64–68(in Lithuanian)
Kirkland GL(1990)Patterns of initial small mammal community change after clearcutting of temperate North American forests.Oikos 59:313–320.doi:10.2307/3545141
Kleemola S,S?derman G(1993)Manual for integrated monitoring,Programme phase 1993–1996.Environmental Report 5.Environmental Data Centre,Helsinki
Kotecky V(2015)Contribution of afforestation subsidies policy to climate change adaptation in the Czech Republic.Land Use Policy 47:112–120.doi:10.1016/j.landusepol.2015.03.014
Lasanta T,Nadal-Romero E,Arnáez J(2015)Managing abandoned farmland to control the impact of re-vegetation on the environment.The state of the art in Europe.Environ Sci Pol 52:99–109.doi:10.1016/j.envsci.2015.05.012
Lithuanian Forest Cover Enlargement Programme(2002)Lietuvos mi?kingumo didinimo programa,patvirtinta Lietuvos Respublikos aplinkos ministro ir Lietuvos Respublikos ?em?s ūkio ministro 2002 m.gruod?io 2 d.?sakymu Nr.616/471
Ludwig JA,Reynolds JF(1988)Statistical Ecology:A Primer on Methods and Computing.Wiley Press,New York
Ma?eikyt? R(2002)Small mammals in the mosaic landscape of eastern Lithuania:species composition,distribution and abundance.Acta Zool Litu 12:381–391.doi:10.1080/13921657.2002.10512528
Panzacchi M,Linnell JD,Melis C,Odden M,Odden J,Gorini L,Andersen R(2010)Effect of land-use on small mammal abundance and diversity in a forest–farmland mosaic landscape in south-eastern Norway.Forest Ecol Manag 259:1536–1545.doi:10.1016/j.foreco.2010.01.030
Pearce J,Venier L(2005)Small mammals as bioindicators of sustainable boreal forest management.Forest Ecol Manag 208:153–175.doi:10.1016/j.foreco.2004.11.024
Raoul F,Pleydell D,Quere JP,Vaniscotte A,Rieffel D,Takahashi K,Bernard N,Wang J,Dobigny T,Galbreath KE,Giraudoux P(2008)Small-mammal assemblage response to deforestation and afforestation in central China.Mammalia 72:320–332.doi:10.1515/mamm.2008.045
Saitoh T,Nakatsu A(1997)The impact of forestry on the small rodent community oh Hokkaido,Japan.Mammal Study 22:27–38 http://doi.org/10.3106/mammalstudy.22.27
Schoonmaker P,McKee A(1988)Species composition and diversity during secondary succession of coniferous forest in the western cascade mountains of Oregon.For Sci 34:960–979
StatSoft Inc(2013)Electronic Statistics Textbook.StatSoft,Tulsa http://www.statsoft.com/textbook/.Accessed 9 Mar 2015
Sullivan TP,Lautenschlager RA,Wagner RG(1999)Clearcutting and burning of northern spruce-fir forests:implications for small mammal communities.J Appl Ecol 36:327–344.doi:10.1046/j.1365-2664.1999.00408.x
Sullivan TP,Sullivan DS,Lindgren PMF(2000)Small mammals and stand structure in young pine,seed-tree,and old-growth forest,southwest Canada.Ecol Appl 10:1367–1383.http://dx.doi.org/10.1890/1051-0761(2000)010[1367:SMASSI]2.0.CO;2
Swanson ME,Franklin JF,Beschta RL,Crisafulli CM,DellaSala DA,Hutto RL,Lindenmayer DB,Swanson FJ(2010)The forgotten stage of forest succession:early-successional ecosystems on forest sites.Front Ecol Environ 9:117–125 http://dx.doi.org/10.1890/090157
Swihart RK,Slade NA(1990)Long-term dynamics of an early successional small mammal community.Am Midl Nat 123:373–382.doi:10.2307/2426565
?inkūnas R,Bal?iauskas L(2006)Small mammal communities in the fragmented landscape in Lithuania.Acta Zool Litu 16:130–136.doi:10.1080/13921657.2006.10512721
Torre I,Diaz M(2004)Small mammal abundance in Mediterranean post-fire habitats:a role for predators?Acta Oecol 25:137–142.doi:10.1016/j.actao.2003.10.007
Tóthmérész B(1993)DivOrd 1.50:a program for diversity ordering.Tiscia 27:33–44
Tóthmérész B(1998)On the characterization of scale-dependent diversity.Abstr Bot 22:149–156
Urban NA,Swihart RK(2011)Small mammal responses to forest management for oak regeneration in southern Indiana.Forest Ecol Manag 261:353–361.doi:10.1016/j.foreco.2010.10.015
Zar JH(1999)Biostatistical analysis.Prentice-Hall,Upper Saddle River
Zwolak R(2009)A meta-analysis of the effects of wildfire,clearcutting,and partial harvest on the abundance of North American small mammals.Forest Ecol Manag 258:539–545.doi:10.1016/j.foreco.2009.05.033
*Correspondence:linasbal@ekoi.lt;linas.balciauskas@gamtostyrimai.lt;linas.balciauskas@gmail.com
Nature Research Centre,Akademijos 2,08412 Vilnius,Lithuania
?The Author(s).2017 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0
International License(http://creativecommons.org/licenses/by/4.0/),which permits unrestricted use,distribution,and
reproduction in any medium,provided you give appropriate credit to the original author(s)and the source,provide a link to the Creative Commons license,and indicate if changes were made.
Received:10 January 2017 Accepted:5 July 2017

- Forest Ecosystems的其它文章
- Development and evaluation of robust tree biomass equations for rubber tree(Hevea brasiliensis)plantations in India
- Climate change effects in the Western Himalayan ecosystems of India:evidence and strategies
- Dead standing pine trees in a boreal forest landscape in the Kalevala National Park,northern Fennoscandia:amount,population characteristics and spatial pattern
- Stand dynamics of the drought-affected floodplain forests of Araguaia River,Brazilian Amazon
- Tree species diversity and utilities in a contracting lowland hillside rainforest fragment in Central Vietnam
- Conspecific negative density dependence in American beech