
Retrograde traff i cking of VMAT2 and its role in protein stability in non-neuronal cells
2016-04-18 11:17:08QiuziWuHongfeiXuWeiWangFeiChangYuJiangYongjianLiu
Qiuzi Wu,Hongfei Xu,Wei Wang,Fei Chang,Yu Jiang,Yongjian Liu,,?
1Department of Physiology,School of Basic Medical Science,Nanjing Medical University,Nanjing,Jiangsu 211166, China;
2Department of Pharmacology&Chemical Biology,University of Pittsburgh School of Medicine,Pittsburgh,USA.
Retrograde traff i cking of VMAT2 and its role in protein stability in non-neuronal cells
Qiuzi Wu1,Δ,Hongfei Xu1,Δ,Wei Wang1,Fei Chang1,Yu Jiang2,Yongjian Liu1,2,?
1Department of Physiology,School of Basic Medical Science,Nanjing Medical University,Nanjing,Jiangsu 211166, China;
2Department of Pharmacology&Chemical Biology,University of Pittsburgh School of Medicine,Pittsburgh,USA.
Increasing evidence suggests that the impaired neuroprotection of vesicular monoamine transporter 2(VMAT2) contributes to the pathogenesis of Parkinson's disease.That has been linked to aberrant subcellular retrograde traff i cking as strongly indicated by recent genomic studies on familial Parkinson's diseases.However,whether VMAT2 function is regulated by retrograde traff i cking is unknown.By using biochemistry and cell biology approaches,we have shown that VMAT2 was stringently localized to the trans-Golgi network and underwent retrograde traff i cking in non-neuronal cells.The transporter also interacted with the key component of retromer, Vps35,biochemically and subcellularly.Using specif i c siRNA,we further showed that Vps35 depletion altered subcellular localization of VMAT2.Moreover,siRNA-mediated Vps35 knockdown also decreased the stability of VMAT2 as demonstrated by the reduced half-life.Thus,our work suggested that altered vesicular traff i cking of VMAT2 may play a vital role in neuroprotection of the transporter as well as in the pathogenesis of Parkinson's disease.
Parkinson's disease,VMAT2,Vps35,retrograde traff i cking
Introduction
Parkinson's disease(PD)is a progressive neurodegenerative disease which commonly occurs in the elderly population.Its pathological changes are mainly embodied in the dopaminergic neuronal loss in the substantia nigra of midbrain.Clinical manifestations are locomotor ataxia,such as rest tremor,bradykinesia, rigidity and postural instability[1-2].Previous studies on PD pathogenesis have been primarily focused on the fi nding of exogenous neurotoxin and related mechanisms leading to neuronal loss[3-4],whereas the endogenous protective mechanism is poorly investigated. Our laboratory previously found that vesicular monoamine transporters(VMATs,SLC18A family)can protect dopaminergic neurons from the toxicity of neurotoxin MPP+[5-6].Thus,it has been hypothesized that attenuated expression or function of VMAT2,the brain isoform of VMATs,is associated with altered detoxif i cation capacity of the transporter and exacerbation of the cell oxidative state.Although there is increasing evidence supporting such reduced neuroprotection of VMAT2 in PD pathogenesis[7],limited molecular and cellular mechanistic studies have been reported.Recently,works on the mechanism of the vesicular positioning and the membrane traff i cking ofthe transport protein itself provide a physiologic basis for understanding its cellular mechanisms of neuroprotection and potential dysfunction.
VMAT2 facilitates a vesicular package of the newly synthesized and recycled monoamine transmitters, like dopamine,into secretory vesicles such as synaptic vesicles(SVs)and large dense core vesicles (LDCVs)[8–10].Since monoamine transmitters are often readily oxidative in the cytoplasm and may produce toxicity against cells,it is of physiologic signif i cance that VMAT2 can protect neurons by transporting these oxidation-prone chemicals into the vesicles,away from sensitive organelles such as mitochondria[11].Thus,the precise vesicular membrane localization of VMAT2 is vital for maintaining such functions as synaptic transmission and detoxif i cation as well.VMAT2 contains 12 transmembrane domains,and its membrane traff i cking and the correct vesicular localization are determined by the cytosolic C-terminal sequence[12-13]. For instance,the sorting of VMAT2 to the newly formed secretory vesicles occurs at the trans-Golgi network (TGN),and this process may be determined by the acidic patch sequence at the distal end of the C terminus[14].In addition,upon exocytosis VMAT2 requires its dileucine motif for internalization from plasma membrane to be sorted to endosomal compartments[15].However,how the intracellular membrane traff i cking of VMAT2 is regulated remains largely elusive.Importantly,many PD related gene mutations may contribute to aberrant membrane traff i cking,especially endosome-to-TGN retrograde traff i cking[16–18], whereas the specif i c affected cargo protein is still unknown.
It is well established that internalized membrane proteins often encounter three fates at the endosomal compartments[19].First,they recycle back to the plasma membrane through recycling endosomes.Secondly, they undergo degradation through the endo-lysosomal pathway.Thirdly,they may traff i c back to the TGN for reuse through a conserved retrograde traff i cking pathway which is mediated by the retromer.Retromer complex is composed of two subcomplexes:one is the cargo-selective Vps trimer,including Vps26,Vps29 and Vps35,and the other is sorting nexin(SNX)dimer, containing SNX1/2 and SNX5/6,which presumably senses membrane curvature changes[20].Recently,it has been reported that Vps35 has a close relationship with the pathogenesis of neurodegenerative diseases,such as PD[16,18]and Alzheimer's disease(AD)[21-22].Moreover,the mutant Vps35D620Nhas been indicated as gainof-function form involved in PD pathogenesis[16,23]. Nonetheless,the functional specif i city of Vps35 within neuronal cells is far from clear and its potential cargo proteins in the dopaminergic cells are poorly investigated.
Here,we report that Vps35 specif i cally interacted with VMAT2 based on biochemical and cell biology analysis.The depletion of Vps35 by its specif i c siRNA disturbed VMAT2 subcellular localization at the TGN. Moreover,depletion of Vps35 altered the protein stability of VMAT2,suggesting that retrograde traff i cking plays a vital role in subcellular localization and functional regulation of VMAT2.
Materials and methods
Cell culture and transfection
All cells were cultured under a standard protocol described by ATCC and in DMEM supplemented with 10%cosmic calf serum(CCS,HyClone)at 37°C and 5%CO2.CHO stable transformants of rat 3Flag-VMAT2 were prepared according to the protocol described previously in our laboratory[9].
For transient overexpression,Lipofectamine 2000 (Life Technologies)were used and transfected cells were harvested and analyzed 24-48 hours later.For siRNA,transfection was also performed using Lipofectamine 2000 according to the manufacturer's protocol.For immunoblotting assays on protein expression analysis,siRNA knockdown cells were incubated with 1.25 nmol/L of each siRNA for 24 hours.
Antibodies
The following antibodies were used in the study: mouse anti-Flag M2,mouse anti-EEA1 and polyclonal anti-IL2α(Sigma),mouse anti-HA.11(Covance),polyclonal sheep anti-TGN46(AbD Serotec),and polyclonal rabbit anti-CI-M6PR and mouse anti-LAMP1 (Abcam).Secondary donkey anti-rabbit IgG Alexa Fluor488,goat anti-mouse IgG Alexa Fluor568,goat anti-rabbit IgG Alexa Fluor568 and goat anti-sheep IgG Alexa Flour 488 were purchased from Life Technologies.
Plasmids,oligonucleotides and reagents
The cDNA fragments in both plasmids of pcDNA3.1-3Flag-VMAT2 and pcDNA3.1-3HA-Vps35 were generated by proof-reading PCR and then inserted to corresponding vectors.The pcDNA3.1-Tac-M was described previously[15].The following reagents were purchased from Sigma-Aldrich:cycloheximide(CHX), NP-40 and saponin.
Immunoprecipitation and protein half-life analysis
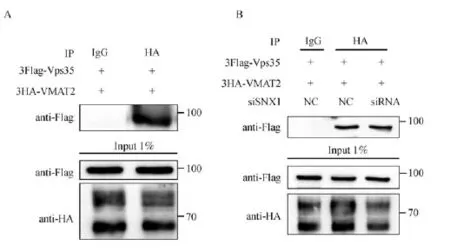
Fig.1 Vps35 specif i cally interacts with VMAT2.A.COS7 cells overexpressing 3Flag-Vps35 and 3HA-VMAT2 were lysed and immunoprecipitated with anti-HA,and analyzed by Western blotting.Input and bound proteins were analyzed by immunoblotting with antibodies against HA and Flag,respectively.The data indicated that Vps35,but not nonspecif i c IgG,interacts with VMAT2.B.COS7 cells overexpressing both 3Flag-Vps35 and 3HA-VMAT2 and cotransfected with SNX1 specif i c siRNAwere lysed and immunoprecipitated with anti-HA.Input and bound proteins were analyzed by blotting with antibodies against HA and Flag,respectively.The data suggested that reduced SNX1 had little inf l uence on the interaction between Vps35 and VMAT2.
Cells were washed with phosphate buffered saline (PBS)and lysed in NP-40 lysis buffer(0.5%NP-40, 150 mmol/L NaCl,50 mmol/L Tris-HCl,pH 7.0 and 5 mmol/L EDTA)supplemented with 1 mmol/L PMSF and 0.1 mmol/L leupeptin for 10 minutes on ice.Cell lysate was centrifuged at 1,600 g and the supernatants were then precleared by incubation for 60 minutes at 4°C with 30 mL protein A/G agarose beads(Thermo Scientif i c Pierce)and centrifugation at 8,000 g for 5 minutes.The precleared lysates were incubated for 2 hours at 4°C with 30mL protein A/G agarose beads bound to polyclonal antibody to tagged protein.After immunoprecipitation,the beads were washed 4 times with wash buffer(0.5%NP-40,150 mmol/L NaCl, 50 mmol/LTris-HCl,pH 7.0 and 5 mmol/L EDTA)and then underwent SDS-PAGE analysis.
For half-life detection,VMAT2 stable transformants were transiently transfected with Vps35 specif i c siRNA. After 24 hours,cells were treated with 100 mg/mL CHX and samples were collected at the time course of 0,2,4, 6,8 and 10 hours.Expression levels of VMAT2 were then analyzed by immunoblotting.Prestained protein standard marker(Thermo Scientif i c)(at.26616)was used for side-labelling Western blot.All the Western blotting analyses were performed utilizing the Tanon 5200 Multi Autoimaging System(Tanon,Shanghai).
Immunof l uorescence
For double immunostaining,cells were plated onto glass coverslips coated with poly-D-lysine and Matrigel (Collaborative Research)as previously described[9],and fi xed with 4%paraformaldehyde in 0.1 mol/L phosphate buffer,pH 7.2.After permeabilization and blocking in PBS containing 2%bovine serum albumin, 1%f i sh skin gelatin,and 0.02%saponin for 1 hour,the cells were incubated with primary antibody in the same buffer for 1.5 hours at room temperature,washed 3 times for 10 minutes each in the same buffer,incubated an additional 1 hour at room temperature with the appropriate secondary antibody at an adequate dilution in the same buffer,and washed again 3 times for 5 minutes each.
RNA interference experiment
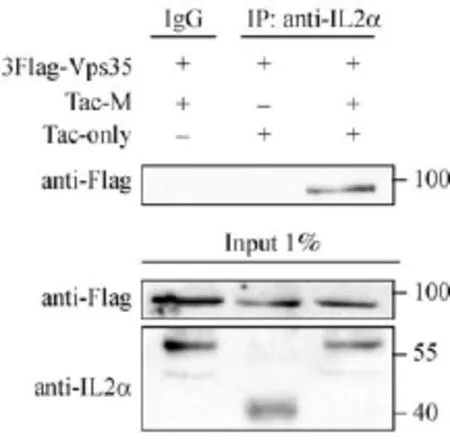
Fig.2 VMAT2 C-terminus is required and suff i cient for the interaction between VMAT2 and Vps35.COS7 cells overexpressing 3Flag-Vps35 and Tac-M or Tac-only were lysed for coimmunoprecipitation with IL-2α polyclonal antibody.Input and bound proteins were analyzed by immunoblotting with antibodies against Tac and Flag,respectively.The data suggested that VMAT2 C-terminus is required and suff i cient for the interaction.
The siRNA oligonucleotides were obtained from GenePharma(Shanghai)and resuspended in doubledistilled water according to the manufacturer's instructions.Thesequences usedfor SNX1 siRNAinterference were5′-CACUCUCAGAAUGGCUUCATT-3′and 5′-GUAGCCUACAAAGUUACAATT-3′.The sequences for Vps35 knockdown study were 5′-GCGUUUGUUUCUUCGAAAUTT-3′,5′-GGAGCAAGUUGUGAAUUGUTT-3′and 5′-GCCUAAAGAAGGCACUAAATT-3′.siRNA transfection was performed using Lipofectamine 2000 according to the manufacturer's protocol(Life Technology).The cells were harvested 48 h after transfectionand analyzed by PAGE with Western blotting or other assays as specif i ed in each experiment.
Statistical analysis
Statistical analysis was performed using the Graph-Pad Prism software(version 5.0,GraphPad Software). For quantitative analysis of immunoblots,the expression level of VMAT2 in half-life studies were quantif i ed by densitometry of the bands between two treatment groups and statistical signif i cance was determined by unpaired t-test as indicated.Results are expressed as the mean±SD if not indicated otherwise.
Results
Vps35 specif i cally interacts with VMAT2
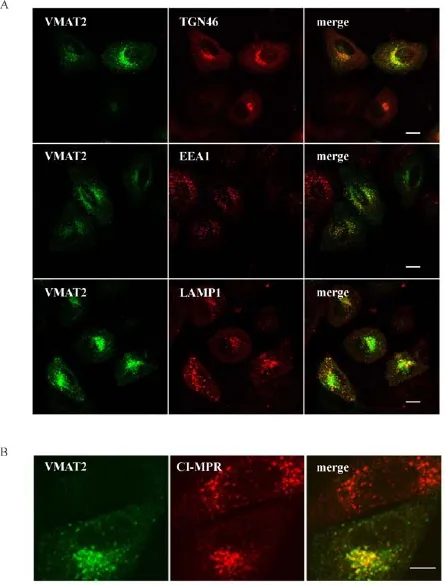
Fig.3 Localization of VMAT2 to TGN A.HeLa cells transiently transfected with plasmids of 3Flag-VMAT2 were immunostained for organelle markers TGN46,EEA1 and LAMP1 followed by confocal microscopic analysis.Pictures showed that VMAT2 was signif i cantly colocalized with TGN46 and partial colocalized with EEA1 and LAMP1.Scale bar,10 mm.B.HeLa cells expressing 3Flag-VMAT2 were double immunostained with Flag antibodies and endogenous CI-MPR followed by confocal microscopic analysis.The data showed that two proteins were signif i cantly co-localized perinuclearly.Scale bar,10 mm.
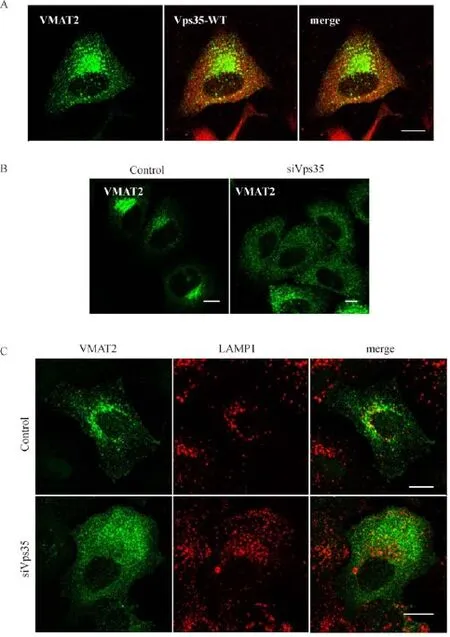
Fig.4 Reduced expression of Vps35 alters the subcellular localization of VMAT2.A.HeLa cells were transiently transfected with two plasmids,3Flag-VMAT2 and 3HA-Vps35,followed by double immunostaining for confocal microscopic analysis.The data showed that wild type Vps35 did not alter the perinuclear staining pattern of VMAT2.Scale bar,10 mm.B and C.HeLa cells expressing 3Flag-VMAT2 were transiently transfected with Vps35 specif i c siRNA and scramble siRNA,and were stained with Flag antibody(B,green),and with LAMP1 conjunction(C,red),and examined by confocal microscopy.The data showed that Vps35 depletion led to mis-targeting of VMAT2 from perinuclear pattern to diffused pattern and increased colocalization with lysosome.Scale bar,10 mm.
Genetic and biochemical analyses have proved Vps35p as the cargo recognition component of the retromer[24].To determine whether Vps35 regulated the membrane traff i cking of VMAT2 through specif i c interaction,we performed immunoprecipitation on COS7 cells co-expressing both Vps35 and VMAT2. Transient transfected cells were lysed in lysis buffer containing 0.5%NP-40 and then HA-tagged VMAT2 was pull down as the bait protein,followed by immunoblotting to determine whether Vps35 can be detected as a binding partner.As shown in Fig.1A, Vps35 was co-immunoprecipitated with overexpressed VMAT2 but not with IgG in the control group, suggesting a specif i c interaction between the two proteins.Importantly,double immunof l uorescence study also showed Vps35 and VMAT2 were partially co-localized in HeLa cells(Fig.4A).Interestingly, when another retromer component,SNX1,was depleted via the specif i c siRNA-mediated knockdown,the interaction between VMAT2 and Vps35 overexpressed in the transient transfected COS7 cells was not signif i cantly altered(Fig.1B),indicating that SNX1 was not required for this functional interaction.Thus, these results suggested that Vps35 specif i cally interacted with VMAT2.
Vps35 binds to the C-terminus of VMAT2
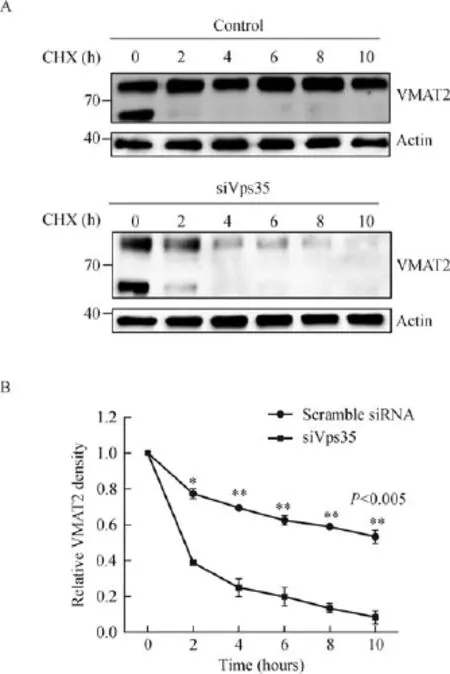
Fig.5 Reduced expression of Vps35 alters the half-life of VMAT2.A.CHO stable transformants of rat 3Flag-VMAT2 were transiently transfected with Vps35 specif i c siRNA.After 24 hours, cells were treated with 100 mg/mL CHX and collected samples at the indicated time points(0,2,4,6,8,and 10 hours).Proteins were analyzed by blotting.Actin is used the loading control.The data suggested that Vps35 depletion caused shorter half-life of VMAT2 compared with that in control group.B.The arbitrary densitometry value in the control group and siRNA group was measured using imaging analysis software Image J.Data are presented as mean± SD,n=3.**P<0.005,unpaired t-test.
To determine how the two proteins interact,we further analyzed domains required for this binding.We used a chimeric protein,Tac-M(the construct was made by fusing an unrelated plasma membrane protein called Tac with the C-terminus of VMAT2)to demonstrate the suff i ciency of the C-terminus of the transporter for the interaction.Again both Flag-Vps35 and Tac-M were overexpressed for co-immunoprecipitation study.As shown in Fig.2,Vps35 interacted with Tac-M but not Tac-only,suggesting that the C-terminus of VMAT2 was required and suff i cient for the interaction.
Vps35 regulates retrograde traff i cking of VMAT2
The interactions described above prompted us to examine potential involvement of Vps35 in subcellular vesicular traff i cking of VMAT2.We f i rst determined whether VMAT2 is subcellularly localized to retrograde traff i cking related organelles such as TGN.Immunofl uorescence assay showed that VMAT2 was signi ficantly colocalized with TGN marker,TGN46,and partially colocalized with endosome(EEA1)and lysosome(LAMP1)(Fig.3A).Importantly,VMAT2 showed extensive colocalization with endogenous cation-independent mannose 6-phosphate receptor(CIM6PR),a classical retrograde traf fi cking cargo as well as a well-known marker for TGN(Fig.3B).Thus,these results clearly indicated that VMAT2 served as a target of retromer,and Vps35 may be involved in regulating its retrograde traf fi cking.To further investigate such possibility,we fi rst showed VMAT2 and Vps35 colocalized in overexpressed cells(Fig.4A).We then determined whether VMAT2 subcellular localization was altered by Vps35 depletion.As shown in immunofl uorescent staining in Fig.4B,Vps35 depletion led to mis-targeting of VMAT2 from a perinuclear pattern to a diffused pattern.More importantly,Vps35 KD cells showed increased co-localization of VMAT2 and LAMP1,suggesting that Vps35 regulated the TGN targeting of VMAT2,possibly keeping it away from lysosome mediated degradation(Fig.4C).
Vps35 depletion decreases the half-life of VMAT2
To determine whether Vps35 plays a role in protecting VMAT2 from degradation,the half-life of VMAT2 in Vps35 KD cells was tested.We transfected Vps35 specif i c siRNA into a CHO stable transformant expressing Flag-VMAT2 cells followed by collecting the samples at an interval of 2 hours after the blocking of protein synthesiswith cycloheximide.The half-life of wildtype VMAT2 is approximately 18-20 hours according to our previous study.Vps35 knockdown caused a signif i cantly decreased half-life compared with the control group,with a dramatica reduction of the VMAT2 level to less than half of the beginning in 2 hours(Fig.5A,B).Of note,as shown in Fig.5A, VMAT2 has mature and immature forms,and the immature form will transform to the mature form over time,nearly 2 hours based on our experiments.These data indicated that Vps35 mediated VMAT2 entering retrograde traff i cking pathway for reuse,leading to an increased'using life'of VMAT2.In conclusion,Vps35 mediated retrograde traff i cking may be important in vesicular targeting of VMAT2,keeping it away from lysosomal degradation while maintaining VMAT2 stability in check.
Discussion
In this paper,we investigated whether retromer complex regulated the membrane traff i cking ofVMAT2 using biochemical and cell biology methods. By interaction with VMAT2,Vps35 not only determined retrograde traff i cking related subcellular localization at TGN but was also involved in the protein stability of the transporter.The study demonstrated that VMAT2 underwent stringent retrograde traff i cking in non-neuronal cells and may be subject to subcellular regulation of retromer complex in neurons.
It has been established that VMATs requires specif i c mechanism for targeting to secretory vesicle[25].Newly synthesized transport proteins may be en route cytoplasm membrane to be recruited to synaptic vesicle, which depends on a stringent internalization signal dileucine motif[26].In non-neuronal cells,internalized VMAT2 is localized to the endosomal compartment to be functional in transport activity mediated detoxif i cation[5].However,VMATs may also contain sorting information for retrograde traff i cking.Our previous work indicated that VMAT1 appeared at perinuclear localization in CHO transformants[27].The work reported here conf i rmed such intracellular organelle association as TGN for VMAT2 with much more relevant markers(Fig.3A).Furthermore,series data supports that such localization can be regulated by retromer protein Vps35.Thus,this retrograde traff i cking related subcellular phenomenon may suggest a molecular mechanism for regulating vesicular targeting and function such as neuroprotection of VMAT2 in neuron.
As a key component of the extremely conserved retromer complex,Vps35 often serves as a cargo recognition protein[28].Its interaction with VMAT2 provides a regulatory connection to the transporter in retrograde traff i cking.With little inf l uence from overexpressed SNX1,another component of retromer,we may conclude that the interaction is rather specif i c. However,the current detection assay heavily relies on an overexpression system for both proteins in COS7 cells.Without detailed interaction on endogenous proteins or other physiologic measurement on isolated proteins,it is still hard to rule out the possibility of other protein(s)involved in the retromer-mediated binding. Moreover,recent studies suggested that mammalian cells seem to have more regulatory crosstalk with other cytosolic machinery such as WASH complex and sorting nexins other than classical SNX subcomplex[29,30].Thus,how VMAT2 is regulated on its retrograde traff i cking and the nature of its regulatory machinery in neurons may still be the motivation of further study.Importantly,recent reports on gain of function Vps35 mutant in familiar PD suggest a role of altered membrane traff i cking in PD pathogenesis.Our work here may provide an insight in potential mechanism underlying PD pathogenesis.
Acknowledgements
This work was supported by the National Nature Science Foundation of China(Grant No.31371436 and No.8157051134)and by the laboratory start-up grant from Nanjing Medical University to Y.Liu.We also thank Dr.Steven Cheng's lab for the assistance with confocal laser microscopy.The funders had no role in study design,data collection and analysis,decision to publish,or preparation of the manuscript.
[1] Shulman JM,De Jager PL,Feany MB.Parkinson's disease: genetics and patho genesis[J].Annu Rev Pathol,2011,6: 193–222.
[2] Thomas,B.and M.F.Beal,Parkinson's disease[J].Hum Mol Genet,2007,R183–94.
[3] Picconi B,Piccoli G,Calabresi P.Synaptic dysfunction in Parkinson's disease[J].Adv Exp Med Biol,2012,970:553–572.
[4] Kim-Han JS,Antenor-Dorsey JA,O'Malley KL.The parkinsonian mimetic,MPP+,specif i cally impairs mitochondrial transport in dopamine axons[J].J Neurosci,2011,31(19): 7212–7221.
[5] Liu Y,Peter D,Roghani A,et al.A cDNA that suppresses MPP +toxicity encodes a vesicular amine transporter[J].Cell,1992, 70(4):539–551.
[6] Liu Y,Edwards RH.The role of vesicular transport proteins in synaptic transmission and neural degeneration[J].Annu Rev Neurosci,1997,20:125–156.
[7] Miller GW,Erickson JD,Perez JT,et al.Immunochemical analysis of vesicular monoamine transporter(VMAT2)protein in Parkinson's disease[J].Exp Neurol,1999,156(1):138–148.
[8] Park JJ,Gondré-Lewis MC,Eiden LE,et al.A distinct trans-Golgi network subcompartment for sorting of synaptic and granule proteins in neurons and neuroendocrine cells[J].J Cell Sci,2011,124(Pt 5):735–744.
[9] Liu Y,Edwards RH.Differential localization of vesicular acetylcholine and monoamine transporters in PC12 cells but not CHO cells[J].J Cell Biol,1997,139(4):907–916.
[10]Liu Y,Schweitzer ES,Nirenberg MJ,et al.Preferential localization of a vesicular monoamine transporter to dense core vesicles in PC12 cells[J].J Cell Biol,1994,127(5):1419–1433.
[11]Lohr KM,Miller GW.VMAT2 and Parkinson's disease: harnessing the dopamine vesicle[J].Expert Rev Neurother, 2014,14(10):1115–1117.
[12]Onoa B,Li H,Gagnon-Bartsch JA,et al.Vesicular monoamine and glutamate transporters select distinct synaptic vesicle recycling pathways[J].J Neurosci,2010,30(23):7917–7927.
[13]Nirenberg MJ,Liu Y,Peter D,et al.The vesicular monoamine transporter 2 is present in small synaptic vesicles and preferentially localizes to large dense core vesicles in ratsolitary tract nuclei[J].Proc Natl Acad Sci U S A,1995,92(19): 8773–8777.
[14]Waites CL,Mehta A,Tan PK,et al.An acidic motif retains vesicular monoamine transporter 2 on large dense core vesicles [J].J Cell Biol,2001,152(6):1159–1168.
[15]Colgan L,Liu H,Huang SY,et al.Dileucine motif is suff i cient for internalization and synaptic vesicle targeting of vesicular acetylcholine transporter[J].Traff i c,2007,8(5):512–522.
[16]Follett J,Norwood SJ,Hamilton NA,et al.The Vps35 D620N mutation linked to Parkinson's disease disrupts the cargosorting function of retromer[J].Traff i c,2014,15(2):230–244.
[17]Zimprich A,Benet-Pagès A,Struhal W,et al.A mutation in VPS35,encoding a subunit of the retromer complex,causes late-onset Parkinson disease[J].Am J Hum Genet,2011,89(1): 168–175.
[18]Vilari?o-Güell C,Wider C,Ross OA,et al.VPS35 mutations in Parkinson disease[J].Am J Hum Genet,2011,89(1):162–167.
[19]Rajendran L,Annaert W.Membrane traff i cking pathways in Alzheimer's disease[J].Traff i c,2012,13(6):759–770.
[20]Carlton J,Bujny M,Peter BJ,et al.Sorting nexin-1 mediates tubular endosome-to-TGN transport through coincidence sensing of high-curvature membranes and 3-phosphoinositides[J]. Curr Biol,2004,14(20):1791–1800.
[21]Wen L,Tang FL,Hong Y,et al.VPS35 haploinsuff i ciency increases Alzheimer's disease neuropathology[J].J Cell Biol, 2011,195(5):765–779.
[22]Follett J,Bugarcic A,Collins BM,et al.Retromer's Role in Endosomal Traff i cking and Impaired function in Neurodegenerative Diseases[J].Curr Protein Pept Sci,2016.
[23]Braschi E,Goyon V,Zunino R,et al.Vps35 mediates vesicle transport between the mitochondria and peroxisomes[J].Curr Biol,2010,20(14):1310–1315.
[24]Nothwehr SF,Ha SA,Bruinsma P.Sorting of yeast membrane proteins into an endosome-to-Golgi pathway involves direct interaction of their cytosolic domains with Vps35p[J].J Cell Biol,2000,151(2):297–310.
[25]Liu Y,Krantz DE,Waites C,et al.Membrane traff i cking of neurotransmitter transporters in the regulation of synaptic transmission[J].Trends Cell Biol,1999,9(9):356–363.
[26]Tan PK,Waites C,Liu Y,et al.A leucine-based motif mediates the endocytosis of vesicular monoamine and acetylcholine transporters[J].J Biol Chem,1998,273(28):17351–17360.
[27]Chen CX,Huang SY,Zhang L,et al.Synaptophysin enhances the neuroprotection of VMAT2 in MPP+-induced toxicity in MN9D cells[J].Neurobiol Dis,2005,19(3):419–426.
[28]Kim E,Lee Y,Lee HJ,et al.Implication of mouse Vps26b-Vps29-Vps35 retromer complex in sortilin traff i cking[J]. Biochem Biophys Res Commun,2010,403(2):167–171.
[29]Seaman MN,Gautreau A,Billadeau DD.Retromer-mediated endosomal protein sorting:all WASHed up[J]!TrendsCell Biol, 2013,23(11):522–528.
[30]Temkin P,Lauffer B,J?ger S,et al.SNX27 mediates retromer tubule entry and endosome-to-plasma membrane traff i cking of signalling receptors[J].Nat Cell Biol,2011,13(6):715–721.
CLINICAL TRIAL REGISTRATION
The Journal requires investigators to register their clinical trials in a public trials registry for publication of reports of clinical trials in the Journal.Information on requirements and acceptable registries is available at www.icmje.org/faq_clinical.html.
ΔThese two authors contributed equally to this work.
?Corresponding author:Prof.Yongjian Liu,Department of Physiology,School of Basic Medical Science,Nanjing Medical University, Nanjing,Jiangsu 211166,China,Tel:+86-25-86862153;Email:yjliu78@njmu.edu.cn.
Received 14 April 2016,Revised 8 May 2016,Accepted 20 May 2016,Epub 16 June 2016
The authors reported no conf l ict of interests.
?2016 by the Journal of Biomedical Research.All rights reserved
10.7555/JBR.30.20160061
 THE JOURNAL OF BIOMEDICAL RESEARCH2016年6期
THE JOURNAL OF BIOMEDICAL RESEARCH2016年6期
- THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Pathology and molecular characterization of recent Leucocytozoon caulleryi cases in layer f l ocks
- Modified methods for isolation of pancreatic stellate cells from human and rodent pancreas
- Protein inhibitor of activated STAT 4(PIAS4)regulates liver fi brosis through modulating SMAD3 activity
- Chronic intermittent hypoxia induces cardiac inf l ammation and dysfunction in a rat obstructive sleep apnea model
- Assessment of atrial electromechanical interval using echocardiography after catheter ablation in patients with persistent atrial f i brillation
- Elevated thyroid stimulating hormone levels are associated with metabolic syndrome in a Chinese community-based population of euthyroid people aged 40 years and older