
Research progress on the anticarcinogenic actions and mechanisms of ellagic acid
2014-03-29 05:27:17HongMeiZhangLeiZhaoHaoLiHaoXuWenWenChenLinTao
Cancer Biology & Medicine 2014年2期
Hong-Mei Zhang, Lei Zhao, Hao Li, Hao Xu, Wen-Wen Chen, Lin Tao
1Medical Sciences Research Center,
2Department of Pharmacy, The Second Af fi liated Hospital of Southeast University, Nanjing 210003, China;
3Nanjing Longyuan Natural Polyphenol Synthesis Institute, Nanjing 210042, China
Research progress on the anticarcinogenic actions and mechanisms of ellagic acid
Hong-Mei Zhang1, Lei Zhao1, Hao Li2, Hao Xu3, Wen-Wen Chen3, Lin Tao3
1Medical Sciences Research Center,
2Department of Pharmacy, The Second Af fi liated Hospital of Southeast University, Nanjing 210003, China;
3Nanjing Longyuan Natural Polyphenol Synthesis Institute, Nanjing 210042, China
Cancer is a leading cause of death worldwide. Cancer treatments by chemotherapeutic agents, surgery, and radiation have not been highly e ff ective in reducing the incidence of cancers and increasing the survival rate of cancer patients. In recent years, plant-derived compounds have aracted considerable aention as alternative cancer remedies for enhancing cancer prevention and treatment because of their low toxicities, low costs, and low side e ff ects. Ellagic acid (EA) is a natural phenolic constituent. Recent in vitro and in vivo experiments have revealed that EA elicits anticarcinogenic effects by inhibiting tumor cell proliferation, inducing apoptosis, breaking DNA binding to carcinogens, blocking virus infection, and disturbing in fl ammation, angiogenesis, and drug-resistance processes required for tumor growth and metastasis.is review enumerates the anticarcinogenic actions and mechanisms of EA. It also discusses future directions on the applications of EA.
Ellagic acid (EA); cancer; mechanism
Introduction
Cancer is a leading cause of death in developed and developing countries1,2. Searching for new anticancer agents from plant sources is a realistic and promising approach which may lead to the discovery of many novel anti-cancer drugs3,4.
Ellagic acid (EA) is a naturally occurring phenolic constituent that is contained in ellagitannins in grapes, nuts, strawberries, black currents, raspberries, green tea, pomegranates, and the stem and bark of Eucalyptus globulus, Eucalyptus maculata, and nuts5.e International Union of Pure and Applied Chemistry name of EA is 2,3,7,8-tetrahydroxy-chromeno[5,4,3-cde]chromene-5,10-dione. The total EA quantities of different fruits and plants are provided in Table 1. EA exerts potent preventive and therapeutic effects against several types of cancers, including colon cancer, breast cancer, prostate cancer, skin cancer, esophageal cancer,and osteogenic sarcoma14,15.e anticarcinogenic properties of EA have drawn increasing aention globally.
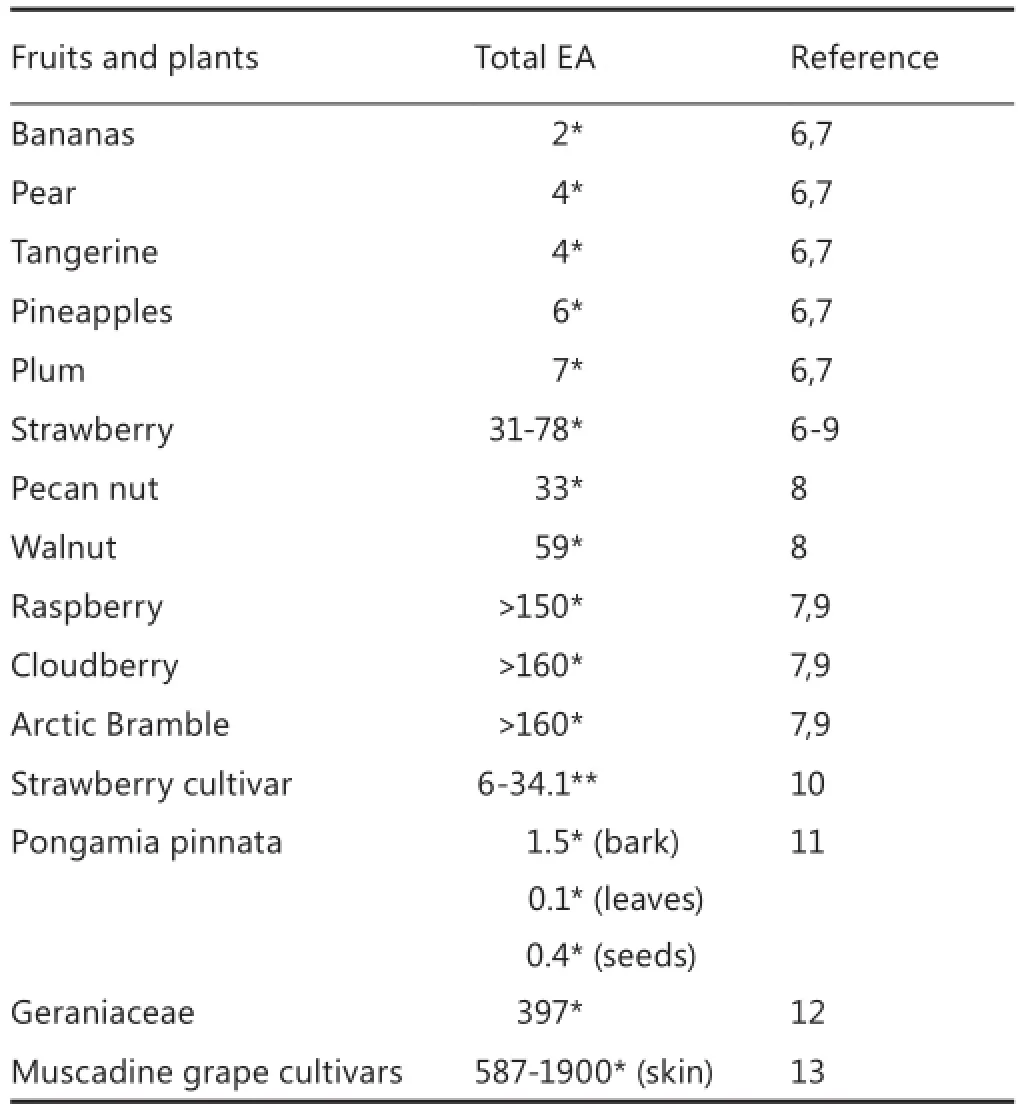
Table 1 Total EA concentration of different plants
Anticarcinogenic e ff ects and mechanisms of EA
Antiproliferative and pro-apoptotic actions and their e ff ects on subcellular signaling pathways
EA has in vitro and in vivo cancer chemopreventive properties. EA treatment is a new and highly e ff ective strategy in reducing cancer carcinogenesis16. EA exerts anticancer e ff ects through its antiproliferative and pro-apoptotic actions, as well as their e ff ects on subcellular signaling pathways.
Han et al.17observed that EA significantly reduces the proliferation and induces the apoptosis of human osteogenic sarcoma (HOS) cells as evidenced by chromosomal DNA degradation and apoptotic body appearance. EA progressively decreased the relative proliferation of the HOS cells in a dosedependent manner, with an IC50value of 6.5 μg/mL. Aside from the progressive, time- and dose-dependent increase in chromosomal DNA degradation, an increase in hypodiploid DNA content and significant time-dependent nuclear fragmentation was also observed.
EA at high concentrations (10-50 mmol/L) can stimulate the apoptosis and completely inhibit the proliferation of the human pancreatic adenocarcinoma cell lines MIA PaCa-2 and PANC-1. EA can accomplish these e ff ects by decreasing nuclear factorkappa B (NF-κB) activity, thereby activating the mitochondrial death pathway, which is associated with loss of mitochondrial membrane potential (Δψm), cytochrome C release, and caspase-3 activation18.
A recent study has indicated that EA at low concentrations (0.5-3 μM) triggers the apoptosis and inhibits the proliferation of the human pancreatic cancer cell lines MIA PaCa-2 and HPAF-II cells. EA elicits similar effects on pancreatic stellate cells, the progenitors of pancreatic cancer desmoplasia. In vivo dietary EA alone can decrease the size and cellularity of a tumor in a subcutaneous xenogramouse model of pancreatic cancer19. Another in vivo study indicated that EA can inhibit pancreatic cancer growth in Balb/C nude mice; this inhibitory effect of EA was associated with the suppression of cell proliferation, activation of caspase-3, and induction of poly (ADP-ribosyl) polymerase cleavage. EA can also inhibit the expression of Bcl-2, cyclin D1, CDK2, and CDK6 while induce the expression of the pro-apoptotic protein Bax in tumor tissues as compared with untreated control tissues20.
Furthermore, EA (10-100 μM) can inhibit the proliferation of ovarian carcinoma ES-2 and PA-1 cells in a dose- and timedependent manner by arresting both cell lines at the G1phase. EA can accomplish these effects by increasing the expression of p53 and Cip1/p21 and decreasing the expression of cyclins D1 and E. EA can also induce caspase-3-mediated apoptosis by increasing the Bax/Bcl-2 ratio, one of the major phenomena that regulate apoptosis, and restore anoikis in both cell lines21.
A previous study22in the human nasopharyngeal carcinoma cell line (NPC-BM1) indicated that EA reduces cell viability. The apoptosis features showed that DNA fragmentation and increased caspase-3 activity are associated with Bcl-2 downregulation. Furthermore, treatment of NPC-BM1 cells can inhibit human telomerase reverse transcriptase and human telomerase-associated protein 1, thereby decreasing telomerase activity.
Vanella et al.16studied the precise molecular mechanisms involved in EA-induced apoptosis in prostate cancer cells. EA produces antiproliferative effects by inhibiting the activation of mammalian target of rapamycin and reducing the intracellular levels of β-catenin in the LNCaP human prostatic cancer cell line. EA also increases the percentage of apoptotic cells by downregulating anti-apoptotic proteins and silencing information regulator 1, human antigen R, and heme oxygenase-1. Furthermore, EA modulates the expression of apoptosis-inducing factor and the activation of caspase-3. Finally, EA increases the expression of the tumor suppressor protein p21.
The protein kinase C (PKC) signaling pathway is critical to cell proliferation, and over activation leads to abnormal tumor growth. The anticarcinogenic action of EA was confirmed after being administered to Dalton’s lymphoma-bearing mice23. EA acts by downregulating PKC, NF-κB, and c-Myc while upregulating transforming growth factor-β1 (TGF-β1). Lymphoma prevention by EA is further supported by the decrease in cell proliferation, cell viability, and ascite fluid accumulation, as well as the increase in the life span of DL mice. A new study from the same research group suggested that EA induces cancer cell death by blocking energy metabolism24.
Breast cancer is the most commonly diagnosed cancer among women worldwide. Two receptor pathways, estrogen receptor and tyrosine kinase receptors, especially the epidermal growth factor receptor family, are drivers of cell proliferation. These pathways are crucial to the development of both primary and recurrent breast cancers. EA not only interacts with and alters the e ff ects of these pathways but also induces cell death (apoptosis and autophagy) by influencing kinase signaling in vitro and in vivo25. Furthermore, these pathways may prevent mammary tumors by suppressing the levels of E2-metabolizing enzymes during early-phase E2 carcinogenesis26.
Prevention of DNA damage generated by oxidative stress and carcinogens
Epidemiological studies and large-scale clinical prevention trials suggested that oxidative stress causes genetic instabilities and functions in the initiation of human cancer.erefore, e ff ective inhibition of endogenous oxidative DNA damage may be a useful prevention strategy27. EA has high effectiveness in preventing oxidative DNA damage both in vitro and in vivo.
EA is a naturally occurring broad spectrum antioxidant.e primary antioxidant mechanism of EA has been aributed to the direct scavenging of free radicals, nitrogen reactive species, and ROS, including hydroxyl radicals, peroxyl radicals, NO2radicals, and peroxynitrite. Other potential protective mechanisms of EA include shielding of DNA from aack and subsequent mutation by its direct association with this macromolecule, inhibition of ROS production, and chelation of metal ions, such as copper28,29.
EA at low doses (1 μM) is substantially effective (nearly 50% inhibition) in preventing dopamine/Cu (II)-mediated oxidatively generated DNA damage30. In EA-treated Chinese hamster lung fibroblast (V79-4) cells, more than 75% of the DPPH radical was scavenged in concentrations of 0.8-100 μg/mL. EA at 4, 20, and 100 μg/mL can inhibit lipid peroxidation by 55%, 79%, and 88%, respectively (IC50value <4.0 μg/mL). The activities of the antioxidant enzymes superoxide dismutase, catalase, and glutathione peroxidase were also signi fi cantly increased in the EA-treated V79-4 cells17.
The cytotoxic and antiproliferative activities of EA against cancer cells do not affect normal cell viability. That is, EA is selectively cytotoxic to carcinoma cells but not to normal cells. A recent study31has indicated that EA has minimal pro-oxidant nature but signi fi cant antioxidant property.
Microarray analysis32revealed that EA modulates several genes. Specifically, EA overexpresses genes involved in DNA repair, such as xeroderma pigmentosum group A complementing protein, DNA ligase III, and DNA excision repair protein, by threefold to eightfold. By contrast, EA downregulates mitogenactivated protein kinase and MAP kinase kinase, which are involved in key cell-signaling pathways.
EA as a chemopreventive agent inhibits carcinogen bioactivation, carcinogen-to-DNA binding, and cancer cell growth33. For example, the formation of O6-methylguanine (O6-meGua) adducts and their persistence are closely linked to esophageal tumor induction in rats. The detection of O6-meGua adducts in the DNA of normal esophageal tissue taken from esophageal cancer patients in China also substantiates the function of methylating nitrosamines in esophageal cancer development. N-nitrosomethylbenzylamine (NMBA) is a procarcinogen that requires metabolic activation to produce its carcinogenic effect. EA exhibits inhibitory effects on NMBA tumorigenesis in the Fischer 344 rat esophagus. When administered in a semi-purified diet at concentrations of 0.4 and 4 g/kg, EA can signi fi cantly reduce (21%-55%) the average number of NBMA-induced esophageal tumors after 20 and 27 weeks of the bioassay34,35. This inhibition is correlated with reductions in the formation of NMBA-induced O6-meGua adducts in esophageal DNA, suggesting that the berries in fl uence NMBA metabolism and reduce DNA damage36.
Prevention of cancer metastasis by inhibiting the angiogenesis, migration, and invasion of cancer cells
Tumor metastasis is a complex cascade that is accompanied by various physiological alterations involved in angiogenesis, matrix metalloproteinase (MMP) upregulation, and extracellular matrix degradation; tumor metastasis allows cancer cells to proliferate and invade blood or lymphatic system, thereby enhancing cancer cell invasion and worsening prognosis37.
Angiogenesis is critical to tumor progression and metastasis. EA demonstrates anti-angiogenic effects both in vitro and in vivo. Sartippour et al.38revealed that an oral administration of pomegranate extract standardized to ellagitannin content to severe combined immunodeficient mice can decrease prostate cancer xenograsize, tumor vessel density, VEGF peptide levels, and hypoxia-inducible factor 1α expression.
EA also exerts anti-angiogenetic effects via the VEGFR-2 signaling pathway in breast cancer15. The structure-based interaction between EA and VEGFR-2 was analyzed. EA can form hydrogen bonds and aromatic interactions within the ATP-binding region of the VEGFR-2 kinase unit and thus significantly inhibit a series of VEGF-induced angiogenesis processes, including proliferation, migration, and tube formation of endothelial cells.
Huang et al.39illustrated a detailed mechanism of EA in angiogenesis. EA demonstrates anti-angiogenic effects by inhibiting MMP-2 activity and secretion, as well as suppressing the tube formation and migration of vascular endothelial cells. Suppressed reversion-inducing cysteine-rich protein with Kazal motifs (RECK) expression was observed in numerous human tumors, including colorectal, breast, pancreas, gastric, hepatocellular, prostate, and non-small cell lung carcinoma.e key action of RECK is to downregulate MMP-2 activity. EA-induced RECK at both mRNA and protein levels associates with the decrease in MMP-2 secretion.
The EA treatment of PANC-1 xenografted mice can inhibitthe expression of the markers of angiogenesis (COX-2, HIF1α, VEGF, VEGFR, IL-6, and IL-8) and metastasis (MMP-2 and MMP-9) in tumor tissues. In addition, EA can significantly inhibit phospho-Akt, Gli1, Gli2, Notch1, Notch3, and Hey1. EA can also reverse epithelial-to-mesenchymal transition by upregulating E-cadherin and downregulating Snail, MMP-2, and MMP-9.e data suggest that EA can inhibit pancreatic cancer growth, angiogenesis, and metastasis by suppressing the Akt, Shh, and Notch pathways20.
In recent studies, EA has shown anti-invasive effects on androgen-independent human (PC-3) and rat (PLS10) prostate cancer cell lines; it also decreases the secretion of MMP-2 from both cells37. The authors further verified that EA significantly reduces the proteolytic activity of collagenase/gelatinase secreted from the PLS-10 cell line. In addition, EA dose dependently inhibits collagenase IV activity. EA reportedly inhibits chemotaxis of the breast cancer cells to stromal cell-derived factor 1α (SDF1α), a chemokine that attracts breast cancer cells to the bone40. Wang et al.41showed that EA can inhibit the growth of hormone-dependent and hormone-refractory prostate cancer cells and inhibit their migration and their chemotaxis toward SDF1α. Moreover, EA can increase the expression of cell adhesion genes and decrease the expression of genes involved in cell cycle control and cell migration. Furthermore, EA can increase several well-known tumor-suppression miRNAs, decrease several oncogenic miRNAs, and inhibit the chemokines receptor type 4/SDF1α chemotaxis axis.
The capability of EA to inhibit the invasion of breast and prostate cancer cells makes this compound a potent and e ff ective treatment for cancer prevention.
Prevention of cancer initiation and progression through anti-in fl ammation
Acute inflammation is a part of the defense response, whereas chronic inflammation can lead to hepatocellular carcinoma (HCC), prostate cancer, colon cancer, breast cancer, and other common forms of cancer. The link between inflammation and cancer is tight. HCC is an in fl ammation-related cancer42because the chronic in fl ammatory state is necessary for the initiation and development of liver cancer. Several studies have shown that chronic infections with hepatitis B virus (HBV) and hepatitis C virus (HCV) are major risk factors for HCC development43-45. Chronic inflammation also affects many cellular pathways, leading to fi brosis and cirrhosis and fi nally hepatocarcinogenesis. Colon cancer is another clear example of the tight link between inflammation and cancer. Inflammatory bowel disease ranks among the top three high-risk conditions for colon cancer.e risk for colorectal cancer increases with the duration and extent of the disease, confirming the active function of inflammation in cancer development. The regular use of nonsteroidal antiinflammatory drugs also lowers the mortality from sporadic colon cancer and results in the regression of adenomas in familial adenomatous polyposis patients46.
Several pro-inflammatory gene products are crucial in suppressing apoptosis, proliferation, angiogenesis, invasion, and metastasis. Among these gene products are TNF and members of its super family, including IL-1α, IL-1β, IL-6, IL-8, IL-18, chemokines, MMP-9, VEGF, COX-2, and 5-LOX. The expression levels these genes are principally regulated by the transcription factor NF-κB. NF-κB mediates innate and adaptive immunity by initiating an inflammatory response to pro-inflammatory signals. NF-κB is constitutively active in most tumors and is induced by carcinogens, tumor promoters, carcinogenic viral proteins (HIV-tat, HIV-nef, HIV-vpr, KHSV, EBV-LMP1, HTLV1-tax, HPV, HCV, and HBV), chemotherapeutic agents, and gamma-irradiation.e persistent activation of NF-κB in tumor cells alters their ability to grow and di ff erentiate. One of the best studied consequences of NF-κB activation is the enhanced survival of cancer cells. The role of persistent inflammation in aiding tumor development has led to the NF-κB family of transcription factors being strongly implicated in promoting cancer47,48. Anti-inflammatory agents that suppress NF-κB or NF-κB-regulated products should have a potential in the prevention and treatment of cancer49.
EA reportedly possesses anti-in fl ammatory properties. Khan et al.50explored the possible chemopreventive mechanism of EA against NF-κB using the 3-D structure and X-ray crystallographic structure of the molecules, as well as molecular docking simulation software. They found that EA shows significant binding affinity with the Rel homology domain of the NF-κB precursor protein p105 with a binding energy of ?7.99 Kcal and an inhibition constant of 1.38 μM. However, EA is not as effective as quercetin and 1-caffeoylquinic acid in inhibiting the target molecule. Rocha et al.40discovered that pomegranate juice or a combination containing EA constituent increases breast cancer cell adhesion and decreases cancer cell migration. They also found that pro-inflammatory cytokines/chemokines are significantly reduced by these treatments. Therefore, these treatments have the potential to decrease inflammation and inhibit cancer progression.
Umesalma et al.51investigated the effect of EA on Wistar albino rats with 1,2-dimethylhydrazine-induced colon cancer.ey found that EA demonstrates anti-in fl ammatory property by downregulating inducible nitric oxide synthase, COX-2, TNF-α, and IL-6 through the inhibition of NF-κB, which is a prompter oftumorigenesis.ey also found that EA exerts chemopreventive e ff ects on colon carcinogenesis. Treatment with EA reduces both TGF-β and IL-6 levels in the LNCaP human prostatic cancer cells16.
The major functions and effective dose of EA on different cancer types are summarized in Table 2.
Other indirect mechanisms involved in EA anticancer actions
Radiosensitizing and counter radioresistance actions
Girdhani et al.52investigated the mechanism of action of di ff erent anticancer and antioxidant agents, including EA, on both normal and cancer cells to develop effective protocols in practical radioprotection and cancer radiotherapy. They found that EA presents cytotoxic e ff ects involving oxidative damage, membrane alteration, and damage to nucleic acid when combined with ionizing radiation in tumor cell lines. Further research53indicated that EA can suppress the radiation-induced activation of receptor tyrosine kinases and NF-κB signaling, can modify cell survival and DNA repair e ffi cacy, and may potentiate ceramide signaling. The radiosensitizing and counter radioresistance mechanisms of EA may provide a new approach to develop an effective treatment for cancer.
Antivirus and liver-/heart-protective actions
The incidence of HCC continues to increase globally. Chronic HBV infections progress through stages of increasing in fl ammation associated with fi brosis and thus result in cirrhosis, which predisposes individuals to HCC. EA blocks the HBV-antigen secretion in HBV-infected hepatocytes and ameliorates the immune tolerance caused by HBeAg during HBV infection in HBeAg-Tg mice without adverse reactions54,55. EA also exhibits anti fi brotic activity in the liver tissue of rats with CCl4-induced liver fibrosis. This result suggests that EA can be used as a therapy for HBV infection and thus reduce the risk for liver tumorigenesis.
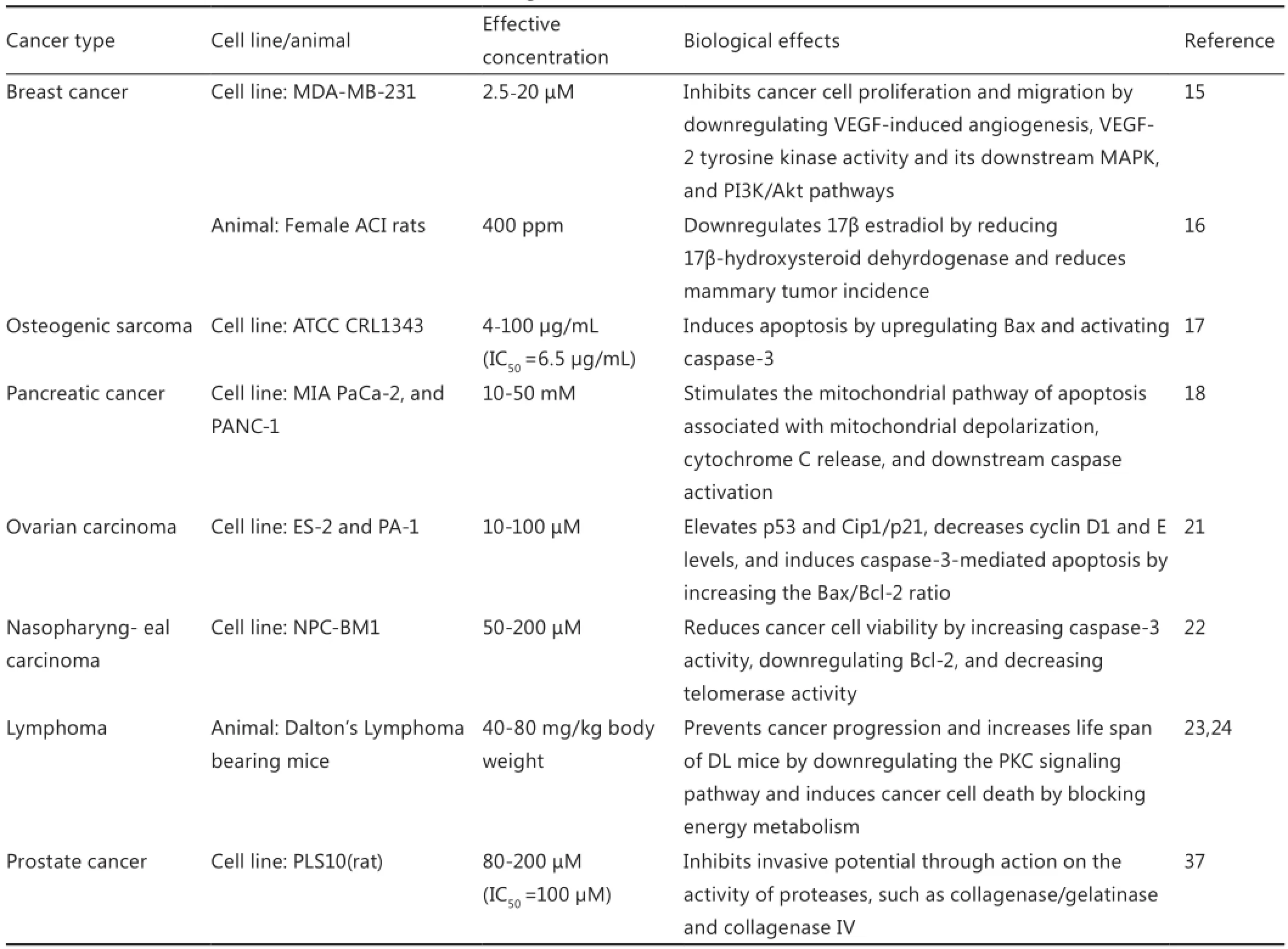
Table 2 Effects of EA in different cancer cell lines or xenografted animals
EA also shows protective effects on liver and heart toxicity induced by cisplatin.erefore, EA may be used in combination with cisplatin in cancer chemotherapy to improve cisplatininduced oxidative stress parameters56.
Inhibiting glutathione S-transferase (GST)-induced drug resistance
GST is a multifunctional detoxification protein that catalyzes the conjugation of glutathione to chemical toxins. GST overexpression in cancer confers resistance to chemotherapeutic agents. Inhibiting GST overexpression has been suggested as an approach to combat GST-induced resistance. EA inactivates the GSTs M1-1, M2-2, and P1-1 in vitro in a time- and concentration-dependent manner.is fi nding suggests that EA can be used as anticancer and chemopreventive agents because of their functions as chemomodulators in GST overexpression in malignancies57.
Current problems and future directions
EA can act through multiple pathways and can be used as a dietary agent for preventing and treating many common forms of cancer.rough the action of human colonic micro fl ora, EA is partially converted into metabolites, including hydroxy-6H-benzopyran-6-one derivatives, primarily urolithin A (UA); then, EA and urolithins enter the circulation58. Recent studies based on in vitro testing have shown preliminary evidence on the antiinflammatory, anticarcinogenic, antiglycative, antioxidative, and antimicrobial effects of urolithins. Although the number of in vivo studies is still limited, their fi ndings on the preventive e ff ects of urolithins on gut and systemic in fl ammation encourage further researches59,60. However, the bioavailability of EA and urolithins is very low. Poor absorption from the gut, rapid metabolism, and lack of transport to the target organs may limit the bioavailability and clinical usefulness of EA and urolithins upon oral administration61. Furthermore, ABC transporters and Phase-II metabolism are involved in cancer cells as a mechanism of cancer resistance against urolithins through their conversion into glucuronide conjugates, which exert low antiproliferative activity60.
To overcome the bioavailability issues, many studies developed drug delivery systems, such as chitosan–glycerol phosphate (C-GP) in situ gelling system for the sustained subcutaneous delivery of EA62, EA-loaded poly (d,l-lactide-coglycolide) nanoparticles for oral administration63, and using a new pH-sensitive polymer [Eudragit P-4135F (P-4135F)] to deliver EA to the lower small intestine in rats64. An increasing number of nanoparticles, liposomes, microemulsions, and polymeric implantable devices are emerging as viable alternatives for delivering therapeutic concentrations of EA into the systemic circulation.e results indicate that the bioavailability of EA has improved65.
EA and its metabolites have preventive and therapeutic potential against human cancers, and advanced drug delivery systems have potential for enhanced bioavailability. However, chemical modifications or more formulations that can bypass their poor oral bioavailability and eliminate hepatic first pass metabolism without compromising patient acceptability must be developed66. Mechanisms to maintain effective therapeutic concentrations in the blood due to the rapid drug metabolism should also be considered.
Acknowledgements
This work was supported in part by grants from The National Science and Technology Foundation for Rural Development in China during the “12th Five-Year-Plan” (Grant No. 2011BAD33B02) ande National Natural Science Foundation of China (Grant No. 81300323).
Con fl ict of interest statement
No potential con fl icts of interest are disclosed.
1. Boyle P, Levin B. eds. World Cancer Report 2008. International Agency for Research on Cancer, 1-260.
3. Ramasamy S, Abdul Wahab N, Zainal Abidin N, Manickam S, Zakaria Z. Growth inhibition of human gynecologic and colon cancer cells by Phyllanthus watsonii through apoptosis induction. PLoS One 2012;7:e34793.
4. Cragg GM, Newman DJ. Plants as a source of anti-cancer agents. J Ethnopharmacol 2005;100:72-79.
5. Girish C, Pradhan SC. Drug development for liver diseases: focus on picroliv, ellagic acid and curcumin. Fundam Clin Pharmacol 2008;22:623-632.
6. Williner MR, Pirovani ME, Güemes DR. Ellagic acid content in strawberries of di ff erent cultivars and ripening stages. J Sci Food Agric 2003;83:842-845.
7. Bakkalba?i E, Mente? O, Artik N. Food ellagitannins-occurrence,e ff ects of processing and storage. Crit Rev Food Sci Nutr 2009;49:283-298.
8. Daniel EM, Krupnick AS, Heur YH, Blinzler JA, Nims RW, Stoner GD. Extraction, stability and, quantitation of ellagic acid in various fruits and nuts. J Food Compost Anal 1989;2:338-349.
9. H?kkinen S, Heinonen M, K?renlampi S, Mykk?nen H, Ruuskanen J, T?rr?nen R. Screening of selected fl avanoids and phenolic acids in 19 berries. Food Research International 1999;32:345-353.
10. Atkinson CJ, Dodds PA, Ford YY, Le Mière J, Taylor JM, Blake PS, et al. E ff ects of cultivar, fruit number and re fl ected photosynthetically active radiation on Fragaria x ananassa productivity and fruit ellagic acid and ascorbic acid concentrations. Ann Bot 2006;97:429-441.
11. Sajid ZI, Anwar F, Shabir G, Rasul G, Alkharfy KM, Gilani AH. Antioxidant, antimicrobial properties and phenolics of di ff erent solvent extracts from bark, leaves and seeds of Pongamia pinnata (L.) Pierre. Molecules 2012;17:3917-3932.
12. Wu QY, Zhou Y, Jin X, Guan Y, Xu M, Liu LF. Chromatographic fi ngerprint and the simultaneous determination of fi ve bioactive components of geranium carolinianum L. water extract by high performance liquid chromatography. Int J Mol Sci 2011;12:8740-8749.
13. Lee, JH, Johnson JV, TalcoST. Identi fi cation of ellagic acid conjugates and other polyphenolics in muscadine grapes by HPLC-ESI-MS. Journal of Agricultural and Food Chemistry 2005;53:6003-6010.
14. Heber D. Multitargeted therapy of cancer by ellagitannins. Cancer Leers 2008;269:262-268.
15. Wang N, Wang ZY, Mo SL, Loo TY, Wang DM, Luo HB, et al. Ellagic acid, a phenolic compound, exerts anti-angiogenesis e ff ects via VEGFR-2 signaling pathway in breast cancer. Breast Cancer Res Treat 2012;134:943-955.
16. Vanella L, Di Giacomo C, Acquaviva R, Barbagallo I, Cardile V, Kim DH, et al. Apoptotic markers in a prostate cancer cell line: E ff ect of ellagic acid. Oncol Rep 2013;30:2804-2810.
17. Han DH, Lee MJ, Kim JH. Antioxidant and apoptosis-inducing activities of ellagic acid. Anticancer research 2006;26:3601-3606.
18. Edderkaoui M, Odinokova I, Ohno I, Gukovsky I, Go VL, Pandol SJ, et al. Ellagic acid induces apoptosis through inhibition of nuclear factor kappa B in pancreatic cancer cells. World J Gastroenterol 2008;14:3672-3680.
19. Edderkaoui M, Lugea A, Hui H, Eibl G, Lu QY, Moro A, et al. Ellagic Acid and Embelin A ff ect Key Cellular Components of Pancreatic Adenocarcinoma, Cancer, and Stellate Cells. Nutr Cancer 2013;65:1232-1244.
20. Zhao M, Tang SN, Marsh JL, Shankar S, Srivastava RK. Ellagic acid inhibits human pancreatic cancer growth in Balb/C nude mice. Cancer Le2013;337:210-217.
21. Chung YC, Lu LC, Tsai MH, Chen YJ, Chen YY, Yao SP, et al.e inhibitory e ff ect of ellagic Acid on cell growth of ovarian carcinoma cells. Evid Based Complement Alternat Med 2013;2013:306705.
22. Huang ST, Wang CY, Yang RC, Chu CJ, Wu HT, Pang JH. Phyllanthus urinaria Increases Apoptosis and Reduces Telomerase Activity in Human Nasopharyngeal Carcinoma Cells. Forsch Komplementmed 2009;16:34-40.
23. Mishra S, Vinayak M. Ellagic acid checks lymphoma promotion via regulation of PKC signaling pathway. Mol Biol Rep 2013;40:1417-1428.
24. Mishra S, Vinayak M. Ellagic Acid Induces Novel and Atypical PKC Isoforms and Promotes Caspase-3 Dependent Apoptosis by Blocking Energy Metabolism. Nutr Cancer 2014;66:675-681.
25. Aiyer HS, Warri AM, WoodDR E, Hilakivi-Clarke L, Clarke R. In fl uence of Berry Polyphenols on Receptor Signaling and Cell-Death Pathways: Implications for Breast Cancer Prevention. J Agric Food Chem 2012;60:5693-5708.
26. Aiyer HS, Gupta RC. Berries and ellagic acid prevent estrogeninduced mammary tumorigenesis by modulating enzymes of estrogen metabolism. Cancer Prev Res (Phila) 2010;3:727-737.
27. Aqil F, Gupta A, Munagala R, Jeyabalan J, Kausar H, Sharma RJ, et al. Antioxidant and antiproliferative activities of anthocyanin/ ellagitannin-enriched extracts from Syzygium cumini L. Nutr Cancer 2012;64:428-438.
28. Priyadarsini KI, Khopde SM, Kumar SS, Mohan H. Free radical studies of ellagic acid, a natural phenolic antioxidant. J Agric Food Chem 2002;50:2200-2206.
29. Gerh?user C, Klimo K, Heiss E, Neumann I, Gamal-Eldeen A, KnauJ, et al. Mechanism-based in vitro screening of potential cancer chemopreventive agents. Mutat Res 2003;523-524:163-172.
30. Spencer WA, Jeyabalan J, Kichambre S, Gupta RC. Oxidatively Generated DNA Damage Following Cu(II)-Catalysis of Dopamine and Related Catecholamine Neurotransmiers and Neurotoxins: Role of Reactive Oxygen Species. Free Radic Biol Med 2011;50:139-147.
31. Weisburg JH, Schuck AG, Reiss SE, Wolf BJ, Fertel SR, Zuckerbraun HL, et al. Ellagic Acid, a Dietary Polyphenol, Selectively Cytotoxic to HSC-2 Oral Carcinoma Cells. Anticancer Res 2013;33:1829-1836.
32. Aiyer HS, Vadhanam MV, Stoyanova R, Caprio GD, Clapper ML, Gupta RC. Dietary Berries and Ellagic Acid Prevent Oxidative DNA Damage and Modulate Expression of DNA Repair Genes. Int J Mol Sci 2008;9:327-341.
33. Whitley AC, Stoner GD, Darby MV, Walle T. Intestinal epithelial cell accumulation of the cancer preventive polyphenol ellagic acidextensive binding to protein and DNA. Biochem. Pharmacol 2003; 66:907-915.
34. Mandal S, Stoner GD. Inhibition of N-nitrosobenzylmethylamine-induced esophageal tumorigenesis in rats by ellagic acid. Carcinogenesis 1990;11:55-61.
35. Siglin JC, Barch DH, Stoner GD. E ff ects of dietary phenethyl isothiocyanate, ellagic acid, sulindac and calcium on the induction and progression of N-nitrosomethylbenzylamineinduced esophageal carcinogenesis in rats. Carcinogenesis 1995;16:1101-1106.
36. Stoner GD, Chen T, Kresty LA, Aziz RM, Reinemann T, Nines R. Protection against esophageal cancer in rodents with lyophilized berries: potential mechanisms. Nutr Cancer 2006;54:33-46.
37. Pitchakarn P, Chewonarin T, Ogawa K, Suzuki S, Asamoto M, Takahashi S, et al. Ellagic Acid inhibits migration and invasion by prostate cancer cell lines. Asian Pac J Cancer Prev 2013;14:2859-2863.
38. Sartippour MR, Seeram NP, Rao JY, Moro A, Harris DM, Henning SM, et al. Ellagitannin-rich pomegranate extract inhibits angiogenesis in prostate cancer in vitro and in vivo. Int J Oncol 2008;32:475-480.
39. Huang ST, Yang RC, Wu HT, Wang CN, Pang JS. Zinc-Chelation Contributes to the Anti-Angiogenic E ff ect of Ellagic Acid on Inhibiting MMP-2 Activity, Cell Migration and Tube Formation. Plos One 2011;6:e18986.
40. Rocha A, Wang L, Penichet M, Martins-Green M. Pomegranate juice and speci fi c components inhibit cell and molecular processes critical for metastasis of breast cancer. Breast Cancer Res Treat 2012;136:647-658.
41. Wang L, Ho J, Glackin C, Martins-Green M. Speci fi c pomegranate juice components as potential inhibitors of prostate cancer metastasis. Transl Oncol 2012;5:344-355.
43. Lim KH, Choi HS, Park YK, Park ES, Shin GC, Kim DH, et al. HBx-induced NF-κB signaling in liver cells is potentially mediated by the ternary complex of HBx with p22-FLIP and NEMO. PLoS One 2013;8:e57331.
44. Branda M, Wands JR. Signal transduction cascades and hepatitis B and C related hepatocellular carcinoma. Hepatology 2006;43:891-902.
45. Kremsdorf D, Soussan P, Paterlini-Brechot P, Brechot C. Hepatitis B virus-related hepatocellular carcinoma: paradigms for viralrelated human carcinogenesis. Oncogene 2006;25:3823-3833.
46. Klampfer L. Cytokines, in fl ammation and colon cancer. Curr Cancer Drug Targets 2011;11:451-464.
47. Mowla SN, Perkins ND, Jat PS. Friend or foe: emerging role of nuclear factor appa-light-chain-enhancer of activated B cells in cell senescence. Onco Targetser 2013;6:1221-1229.
48. Arsura M, Cavin LG. Nuclear factor-kappaB and liver carcinogenesis. Cancer Le2005;229:157-169.
49. Aggarwal BB, Shishodia S, Sandur SK, Pandey MK, Sethi G. In fl ammation and cancer: how hot is the link? Biochem Pharmacol 2006;72:1605-1621.
50. Khan MK, Ansari IA, Khan MS. Dietary phytochemicals as potent chemotherapeutic agents against breast cancer: Inhibition of NF-κB pathway via molecular interactions in rel homology domain of its precursor protein p105. Pharmacogn Mag 2013;9:51-57.
51. Umesalma S, Sudhandiran G. Di ff erential inhibitory e ff ects of the polyphenol ellagic acid on in fl ammatory mediators NF-kappaB, iNOS, COX-2, TNF-alpha, and IL-6 in 1,2-dimethylhydrazineinduced rat colon carcinogenesis. Basic Clin Pharmacol Toxicol 2010;107:650-655.
53. Nambiar D, Rajamani P, Singh RP. E ff ects of phytochemicals on ionization radiation-mediated carcinogenesis and cancer therapy. Mutat Res 2011;728:139-157.
54. Shin MS, Kang EH, Lee YI. A fl avonoid from medicinal plants blocks hepatitis B virus-e antigen secretion in HBV-infected hepatocytes. Antiviral Res 2005;67:163-168.
55. Kang EH, Kown TY, Oh GT, Park WF, Park SI, Park SK, et al.e fl avonoid ellagic acid from a medicinal herb inhibits host immune tolerance induced by the hepatitis B virus-e antigen. Antiviral Res 2006;72:100-106.
56. Yüce A, Ate??ahin A, Ceriba?i AO, Aksakal M. Ellagic acid prevents cisplatin-induced oxidative stress in liver and heart tissue of rats. Basic Clin Pharmacol Toxicol 2007;101:345-349.
57. Hayeshi R, Mutingwende I, Mavengere W, Masiyanise V, Mukanganyama S.e inhibition of human glutathione S-transferases activity by plant polyphenolic compounds ellagic acid and curcumin. Food Chem Toxicol 2007;45:286-295.
58. Vicinanza R, Zhang Y, Henning SM, Heber D. Pomegranate juice metabolites, Ellagic acid and urolithin A, synergistically inhibit androgen-independent prostate cancer cell growth via distinct e ff ects on cell cycle control and apoptosis. Evid Based Complement Alternat Med 2013;2013:247504.
59. González-Sarrías A, Miguel V, Merino G, Lucas R, Morales JC, Tomás-Barberán F, et al.e gut microbiota ellagic acid-derived metabolite urolithin A and its sulfate conjugate are substrates for the drug e ffl ux transporter breast cancer resistance protein (ABCG2/BCRP). J Agric Food Chem 2013;61:4352-4359.
60. González-Sarrías A, Giménez-Bastida JA, Nú?ez-Sánchez MA, Larrosa M, García-Conesa MT, Tomás-Barberán FA, et al. Phase-II metabolism limits the antiproliferative activity of urolithins in human colon cancer cells. Eur J Nutr 2014;53:853-864.
61. Bell C, Hawthorne S. Ellagic acid, pomegranate and prostate cancer -- a mini review. J Pharm Pharmacol 2008;60:139-144.
62. Sharma G, Italia JL, Sonaje K, Tikoo K, Ravi Kumar MN. Biodegradable in situ gelling system for subcutaneous administration of ellagic acid and ellagic acid loaded nanoparticles: evaluation of their antioxidant potential against cyclosporine induced nephrotoxicity in rats. J Control Release 2007;118:27-37.
63. Bala I, Bhardwaj V, Hariharan S, Kharade SV, Roy N, Ravi Kumar MN. Sustained release nanoparticulate formulation containing antioxidant-ellagic acid as potential prophylaxis system for oral administration. J Drug Target 2006;14:27-34.
64. Jeong YI, Prasad Yv R, Ohno T, Yoshikawa Y, Shibata N, Kato S, et al. Application of Eudragit P-4135F for the delivery of ellagic acid to the rat lower small intestine. J Pharm Pharmacol 2001;53:1079-1085.
65. Bansal SS, Goel M, Aqil F, Vadhanam MV, Gupta RC. Advanced drug delivery systems of curcumin for cancer chemoprevention. Cancer Prev Res (Phila) 2011;4:1158-1171.
66. Espín JC, Larrosa M, García-Conesa MT, Tomás-Barberán F. Biological signi fi cance of urolithins, the gut microbial ellagic Acidderived metabolites: the evidence so far. Evid Based Complement Alternat Med 2013;2013:270418.
Cite this article as:Zhang HM, Zhao L, Li H, Xu H, Chen WW, Tao L. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Biol Med 2014;11:92-100. doi: 10.7497/ j.issn.2095-3941.2014.02.004
Hong-Mei Zhang
E-mail: zhmnj2008@aliyun.com
Received March 4, 2014; accepted May 3, 2014. Available at www.cancerbiomed.org
Copyright ? 2014 by Cancer Biology & Medicine
 Cancer Biology & Medicine2014年2期
Cancer Biology & Medicine2014年2期
- Cancer Biology & Medicine的其它文章
- Polymeric nanocomposites loaded with fluoridated hydroxyapatite Ln3+ (Ln = Eu or Tb)/iron oxide for magnetic targeted cellular imaging
- An unusual case of aggressive systemic mastocytosis mimicking hepatic cirrhosis
- Spindle cell carcinoma of the breast as complex cystic lesion: a case report
- Effects of postmastectomy radiotherapy on prognosis in different tumor stages of breast cancer patients with positive axillary lymph nodes
- Clinico-pathological signi fi cance of extra-nodal spread in special types of breast cancer
- Incidence and mortality of female breast cancer in the Asia-Paci fi c region