
Evolutionary importance of the relationship between cytogeography and climate: New insights on creosote bushes from North and South America
2022-10-12 06:48:40RominVidlRussellMrinTdeyRomnUrfusovTomUrfusCintiPolSouto
植物多樣性 2022年5期
Romin Vidl-Russell , Mrin Tdey , Romn Urfusov′ , Tom′ˇs Urfus ,Cinti Pol Souto ,*
a Laboratorio Ecotono- INIBIOMA (CONICET-UNComahue), Quintral,1250, Bariloche, Río Negro, Argentina
b Department of Botany, Faculty of Science, Charles University, Ben′atsk′a 2,128 00, Prague, Czech Republic
A B S T R A C T Relationships between genome size and environmental variables suggest that DNA content might be adaptive and of evolutionary importance in plants. The genus Larrea provides an interesting system to test this hypothesis, since it shows both intra- and interspecific variation in genome size. Larrea has an amphitropical distribution in North and South American deserts, where it is most speciose. Larrea tridentata in North America shows a gradient of increasing autopolyploidy;while three of the four studied South American species are diploids, Larrea divaricata, Larrea nitida, Larrea ameghinoi, and the fourth is an allopolyploid, Larrea cuneifolia. We downloaded available focal species’ georeferenced records from seven data reservoirs. We used these records to extract biologically relevant environmental variables from WorldClim at 30 arc seconds scale, to have a broad characterization of the variable climatic conditions of both regions, and a climatic envelope for each species. We estimated relative DNA content index and relative monoploid genome values, by flow cytometry, of four most abundant Larrea species throughout their respective ranges. Then we winnow the bioclimatic dataset down to uncorrelated variables and sampled locales,to analyse the degree of association between both intra-and interspecific relative DNA content and climatic variables that are functionally relevant in arid environments using Pearson correlations, general linear and mixed effects models. Within the genus Larrea, relative DNA content increases with rising temperature and decreases with rising precipitation. At the intraspecific level, all four species show relative DNA content variation across climatic conditions. Larrea is a genus that shows genome size variation correlated with climate. Our results are also consistent with the hypothesis that extreme environmental pressures may have facilitated repeated whole genome duplication events in North America, while in South America, reticulate evolution, as allopolyploidization, and speciation might have been climate-dependent since the Oligocene.
Keywords:DNA content Drylands Genome size Jarilla Larrea Zygophyllaceae
1. Introduction
Many interesting relationships between DNA content and infraspecific environmental range variables have been described(Grime and Mowforth, 1982; Bennett, 1987; Price, 1988; Poggio and Naranjo, 1990; Bromham et al., 2015). All these reports suggest that plant genome size may be predictable, adaptive, and of evolutionary significance(Poggio et al.,1998;Soltis et al.,2007;Te Beest et al., 2012). Increasing relative DNA content by polyploidization has proven to be an important mechanism of speciation in angiosperms and is a key driver of diversity(De Bodt et al.,2005; Soltis et al., 2009; Tank et al., 2015; Soltis and Soltis, 2016;Landis et al.,2018).Autopolyploid collections are usually treated as multiple copies of a single cytotype within a species, while allopolyploids arise via hybridization between species and are typically viewed as different species. Disentangling the cytological characterization of species across their entire geographic range is important to understand patterns of cytotype formation,establishment and migration, and to uncover previously undetected cytotype variation that can yield better estimates of diversification(Soltis et al.,2007,2010;Severns and Liston,2008;Laport and Ng, 2017).
Drylands phylogeography theory in the Western Hemisphere was developed mostly in North American deserts, but South American deserts were a key centre for the emergence of Neotropical lineages and have been understudied. Some of these lineages later migrated northwards and diversified in the Northern Hemisphere. Moreover, arid land vegetation evolved in the temperate Neotropics since the Oligocene(Burnham and Graham,1999), encompassing heterogeneous climate and topography differing from their current distribution. The genusLarreaCav.(Zygophyllaceae), commonly known as creosote bush, is amphitropically distributed, with species separated spatially by areas that are not bridged by shared vegetation type or climatic regimes.AncestralLarreaspecies is thought to have been South American,and reached North America through long distance dispersal and species have diverged allopatrically since the dispersal events(Barbour,1969; Raven,1972; Lia et al., 2001). The genus exhibits both intra- and interspecific variation in DNA content (Hunziker et al., 1972; Laport et al., 2012), and grows in arid and semiarid regions of North and South America. Phylogeographic studies suggest establishment of polyploid populations of the North American species accompanying its rapid spread (Laport et al.,2012).
Due to the large latitudinal span of the genusLarrea(20-35°N in Mexico and United States, and 22-46°S in Monte Desert of Argentina, plus scattered sites in Chile, Bolivia and Peru), environmental selective pressures should differentially impact species inhabiting different portions of the climatic range.In general,Larreastraddles the 200-mm annual isohyet (Ezcurra et al., 1991), but among North American deserts, precipitation seasonality varies from summer rains, regularly distributed monthly rains to mainly winter rainfall, coupled with an increasing aridity gradient from east to west(Chihuahua desert->Sonora desert->Mojave desert,respectively) (Yang, 1970). Meanwhile, although mean annual precipitation is relatively constant in the Monte Desert, its distribution changes, in a latitudinal fashion, from monsoon-type summer rains in northern Argentina to a winter rainfall peak near the Patagonian Andes, and to a more regular monthly distribution along the southern Atlantic coast (Paruelo et al.,1998; Roig et al.,2009). These environmental characteristics provide a good model for investigating correlations between genome size and climatic variables, which can shed some light on drylands phylogeography in the Americas.
Our goals are to investigate the adaptive potential of DNA content within the genusLarreaand among fourLarreaspecies, by relating variation in relative DNA content to climatic variables throughout their collective distributions in North and South America. We ask the following specific questions: (1) Are there differences in relative DNA content measured by flow cytometry and relative monoploid genome value between species of the genusLarreaacross their collective distributions?(2)Are there differences in climatic ranges amongLarreaspecies?(3)Are climatic variables related to relative DNA content variation in the genusLarrea? We predict that relative DNA content will be higher in harsher(i.e.drier and hotter)climatic conditions,since changes in DNA content may influence an organism's ecological tolerance or increase ecological amplitude or facilitate the acquisition of new genetic functions.Finally,we revisit the idea of integrating estimates of DNA content,measured using modern techniques of flow cytometry, with fine scale, functionally relevant climatic variables, and discuss the evolutionary importance of hybridization and polyploidization within the genus as potential drivers of diversification.
2. Materials and methods
2.1. Study system
We sampled four out of fiveLarreaspecies throughout their respective ranges, leaving outLarrea ameghinoiSpeg., which is an extremely rare species,restricted to few localities in Chubut and Rio Negro provinces in Argentina. The only North American species,Larrea tridentata(DC)Coville has multiple cytotypes,whose ploidy level increase across an aridity gradient; with diploid individuals(2n= 26) occurring in the Chihuahua Desert, tetraploids(2n=4x=52)in the Sonora Desert and hexaploids(2n=6x=78)in the Mojave Desert. Although the taxonomic status of autopolyploids and their influence in biodiversity levels remains a subject of active debate in the literature (Soltis et al., 2007); the differentL. tridentatacytotypes have historically been considered as representing a single species (Yang,1970; Hunziker et al.,1972; Laport et al., 2013). We follow this precedent in our study and consider ploidal variation as representing intra-specific DNA content variation.Interestingly,the 4xand 6xcytotypes are found interdigitated in the field in Sonora and Mojave deserts (Laport and Ramsey,2015). The South AmericanL. nitidaCav., andL. divaricataCav., are diploids (2n= 26), whileL. cuneifoliaCav. is an allotetraploid(2n= 4x= 52) (Hunziker et al.,1972) (Fig.1).
2.2. Field collections
The genusLarreais more speciose in the Monte Desert phytogeographic region in Argentina, which resembles the North American warm deserts, being a heterogeneous mosaic of landscapes,from open sand dune habitats to shrublands dominated by creosote bush. In South America,12 localities were sampled, representing three species,L. cuneifolia,L. divaricata, andL. nitida,totalling 106 individuals collected. Meanwhile, 14 individuals ofL. tridentatawere sampled from three localities, representing the three North American deserts (Chihuahua, Sonora and Mojave Deserts).Two to five individuals per locality were sampled and kept in silica gel until flow cytometry analysis (Table 1). Voucher specimens are deposited in the BCRU herbarium(Bariloche,Argentina).

Table 1 Sampled localities,GPS coordinates and elevation,average relative nuclear DNA content ratio(RCR)and relative monoploid genome value(Cx),along the range of four Larrea species: L. cuneifolia (LC), L. divaricata (LD), L. nitida (LN), and L. tridentata (LT). Number of sampled individuals in brackets. Bioclimatic variables: BIO1: Annual mean temperature (°C), BIO14: Precipitation of the driest month (mm), and Solar radiation: Mean solar radiation (W/m2).
2.3. Flow cytometry and DNA content
We measured relative nuclear DNA content by flow cytometry using the AT-selective fluorochrome 4′, 6′-diamidino-2 phenylindole (DAPI) assay following a simplified two-step protocol(Doleˇzel et al., 2007). We usedSolanum pseudocapsicumL.(2C = 2.61 pg) (Temsch et al., 2010) as primary internal standard,but in some cases peaks of standard and sample overlapped.Therefore, in those cases we employedBellis perennisL.(2C=3.38 pg)(Sch¨onswetter et al.,2007)and we derived genome size ofBellisdirectly from simultaneous analysis withSolanum.We further express relative DNA content values as ratios toSolanum.We chopped leaf tissue together with the standard using a sharp razor blade in a Petri dish containing Otto I buffer(0.1 M citric acid,0.5%Tween 20).We filtered the suspension and incubated for 5 min at room temperature. After incubation, we added 1 ml of Otto II buffer (0.4 M Na2HPO412H2O), along with 2-mercaptoethanol(2 μg/ml) and DAPI (4 μg/ml). All samples were analysed with a Partec CyFlow Space instrument (Partec GmbH, Münster, Germany),equipped with a UV LED chip as the excitation source.Flow cytometry histograms were evaluated using the Partec FloMax software (Partec GmbH). Measurements with coefficient of variation (CV) values, for standard and sample peaks, below 5% were accepted. The extent of the total variation in intraspecific relative DNA content index was calculated as a percentage of the difference between its highest and lowest values, and was expressed as % of the minimum. Also the monoploid genome value (Cx) relative equivalent was calculated by dividing the relative DNA content of each sample by the cytotype ploidy level to test the role of polyploidy in the diversification of the genus.
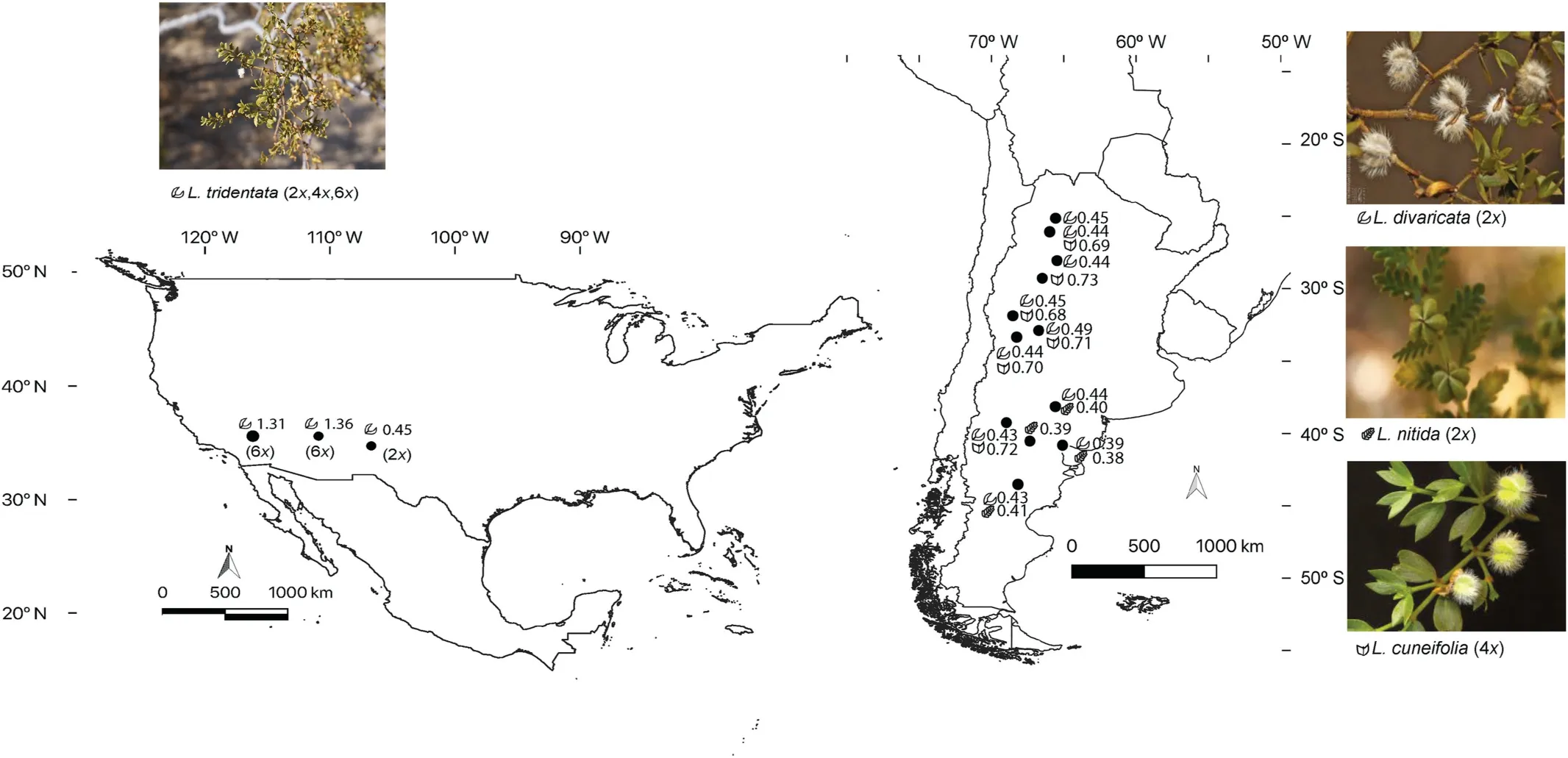
Fig.1. Map of sampled localities of four Larrea species:L.divaricata,L. cuneifolia,L. nitida,L.tridentata in deserts of North and South America.Including average local relative DNA content ratio (RCR) and sketches of leaf morphology.
2.4. Climatic variables
For broad characterization of the variable climatic conditions of both North and South American deserts, we looked for available occurrence data of focal species in seven data reservoirs including herbaria (MO, NY, ARIZ, COLO, SI) and online repositories (GBIF:gbif.org, Mincyt: datos.mincyt.gob.ar/#/biologicos). We then excluded repeated locations or doubtful coordinates. This curated occurrence data set(399 localities),including our sampled localities(15 sites), represents registered GPS-locations for each occurring specie, including latitude, longitude and elevation. For these 399 records we extracted local climate variables from WorldClim 1.4, a set of 19 global bioclimatic variables with a spatial resolution of 30 arc seconds (Hijmans et al., 2005), (http://www.worldclim.org/)(Supplementary material S3). For further analyses we winnowed down the dataset to a reduced set of temperature and precipitation proxies that are functionally relevant in arid environments keeping uncorrelated bioclimatic variables.The climatic matrix included the following variables: BIO1: Annual mean temperature, and BIO14:Precipitation of the driest month and solar radiation.Solar radiation data for South American localities was generated with the ClimateSA v.1.0 software package, available at tinyurl.com/ClimateSA(Hamann et al., 2013), and for North American localities with ClimateNA available at climatena.ca(Wang et al.,2016).Solar radiation is particularly relevant since it has been suggested to have influenced the evolution of leaf orientation forLarreaspecies (Ezcurra et al., 1991). We used this matrix to characterize the climatic envelope for each species,and a subset of the climatic values for the 15 sampled localities were correlated with relative DNA content data.
2.5. Statistical analyses
We tested for differences among species in relative DNA content ratio (RCR) and monoploid genome value (Cx), via Kruskal-Wallis one-way analysis of variance on ranks models and Tukey'sposthoctests. To study the degree of ecological divergence among species,we used boxplots for the climatic variables for each species to graphically depict observed differences among species.As many of the variables demonstrated a non-normal distribution, we used Kruskal-Wallis tests,followed by a nonparametricpost-hoctest,to check the significance of differences between species, using all pairwise multiple comparison procedure (Dunn's Method). To investigate the relationship between climate and relative DNA content, we performed Pearson's correlations and linear mixed effect models, using species identity as random factor. Statistical analyses were carried out in R 3.4.4 (R Development Core Team,2017), using above mentioned functions and, when appropriate,packages nlme and ggplot for graphic representation of the results.
3. Results
3.1. Flow cytometry
In this study we sampled for flow cytometry two diploid species, one species comprising diploids, autotretraploids and autohexaploids, and one allotetraploid species within the genusLarrea(Table 1, Supplementary material S1). As expected,L. tridentatashowed different relative DNA content ratios (RCR)that represent different cytotypes. Samples from New Mexico(Chihuahua desert), have a diploid cytotype, and showed an average RCR (=sample:standard fluorescence ratio (±s.d.)) of 0.445 ± 0.004; while the other two North American populations showed greater RCR, as they are known to be polyploids: Tucson(6x, Sonora desert) yields RCR of 1.360 ± 0.013, and Baker (6x,Mojave Desert) yields RCR of 1.315 (Table 1, Supplementary material S1). DiploidL. tridentataand the South American diploidL. divaricatahave similar RCR and relative monoploid genome values (Cx) (Table 1, Supplementary material S1).L.divaricatahas a mean RCR of 0.433±0.027,the total variation for this species was 38.98% of the minimum value, with minimum RCR in the South(Rio Negro,0.392±0.005)and maximum in the North (Cafayate, 0.450 ± 0.008).L. cuneifoliais the South American species with highest RCR and lower Cx as it is an allotetraploid. Across its geographic distribution, this species has an average RCR of 0.702 ± 0.044 and Cx of 0.175 ± 0.011 (measured withB.perennis,and recalculated forS.pseudocapsicumstandard),total variation for this species was 37.36% of the minimum value.L.nitidais the diploid species with the lowest RCR(0.397±0.013),that varied from 0.383 ± 0.004 in southeast Atlantic coast (Las Grutas) to 0.407 ± 0.004 in the South (Madryn), with total genome size variation of 12.35% of the minimum value (Table 1,Supplementary material S1).
3.2. Climatic envelopes
Data based on the 399 records matrix obtained from online repositories and sampled localities shows thatLarreaspecies grow at statistically different temperature and precipitation ranges(Fig.2,Supplementary material S2,S3).L.divaricataandL.cuneifoliagrow under similar conditions in terms of mean annual temperature (15.01 and 15.33°C), whileL. nitidagrows at lower temperatures (11.80°C) andL. tridentatain North America grows at much higher temperatures (18.22°C) (Fig. 2). In terms of precipitation,L.divaricata(8.36 mm)has a wider range than the other species.L.cuneifolia(7.09 mm)differs fromL.nitidaandL.tridentata,the latter is the most different since it grows in sites with very low precipitation levels(4.2 mm).WhileL.nitidagrows in higher precipitation sites(9.8 mm)(Fig.2).In terms of mean solar radiationL.divaricataandL. cuneifoliagrow under similar conditions (16,624 and 17,550 W/m2), whileL. nitidagrows at lower radiation (15,415 W/m2), andL. tridentatagrows at much higher solar radiation(19,521 W/m2) (Fig. 2).
3.3. Relative DNA content and climatic gradients
When the four studied species are analysed together there is a general pattern suggesting that DNA content increases in warmer and drier climates, with higher solar radiation (Fig. 3, Table 2,Supplementary material S2). All three examined variables are significantly correlated with the relative DNA content for the genus,although correlation is around 0.5. Moreover, within each species some environmental variables showed apparent covariation with relative DNA content ratio (RCR), but they were not significantly correlated, probably due to the low sample size (Table 2). InL.cuneifolia,the South American allotetraploid,RCR decreases with elevation and increases with summer precipitation (southward),with higher DNA content in sites with lower precipitation all year round,i.e.less precipitation seasonality.With higher solar radiation RCR inL. cuneifoliadecreases. InL. divaricataRCR increases northward where summer precipitation decreases(i.e.less precipitation→higher DNA content). RCR inL. nitidaincreases with elevation and decreases with summer temperature and precipitation,and,as withL. cuneifolia, increases when precipitation has more seasonality, i.e. restricted to certain season of the year. The North American species,L. tridentata, show lower RCR eastward and it decreases with summer precipitation, but increases with summer temperature, and precipitation seasonality (Table 2).
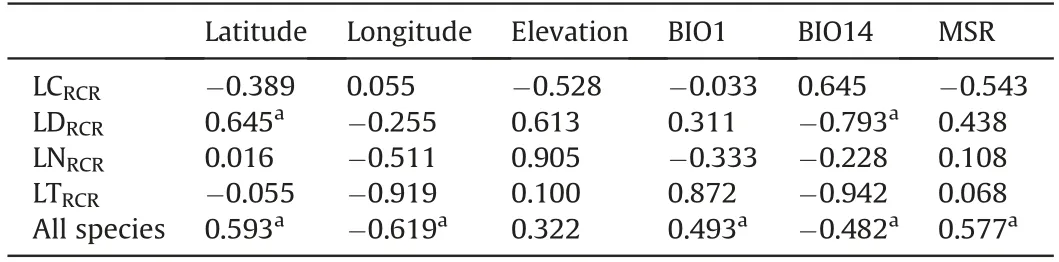
Table 2 Pearson's correlations between each sampled species' relative DNA content ratio(RCR)and geographical and climatic variables:BIO1:Annual mean temperature(°C),BIO14: Precipitation of the driest month (mm), and MSR: Mean solar radiation(W/m2),in four Larrea species:L.cuneifolia(LC),L.divaricata(LD),L.nitida(LN),and L. tridentata (LT).
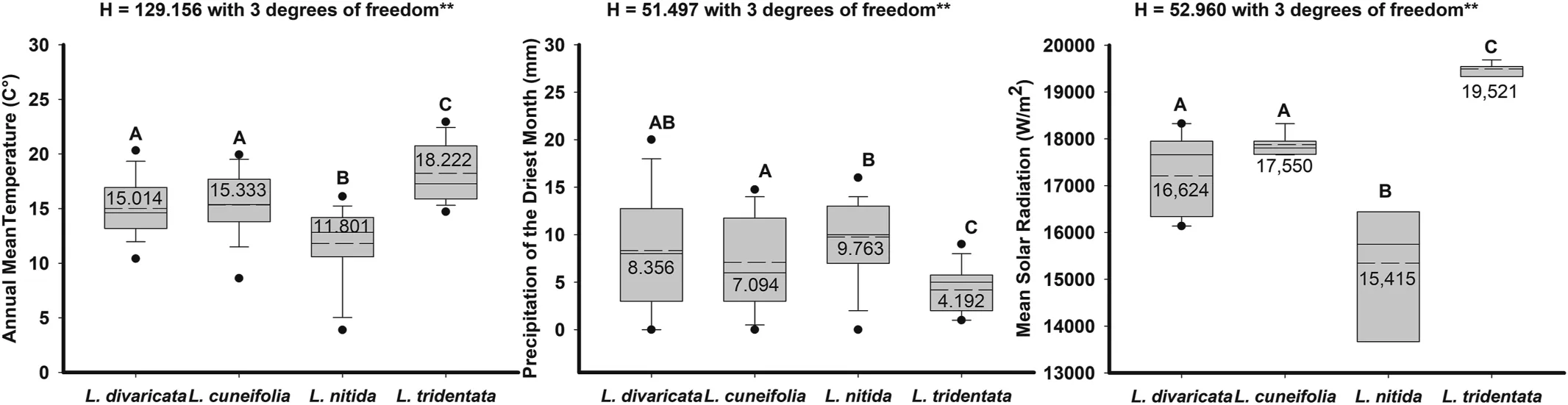
Fig. 2. Box plot of bioclimatic variables extracted from WorldClim using occurrence records of focal species obtained from herbaria, online repositories and our field collections:Annual mean temperature(BIO1),Precipitation of the driest month(BIO14),and Mean solar radiation in four Larrea species.Central complete and dashed lines in the square are the mean and median values,respectively,the box represents the standard deviation,and whiskers are 95%confidence intervals.Including results of Kruskal-Wallis One Way Analysis of Variance on Ranks, different letters represent All Pairwise Multiple Comparison Procedures (Dunn's Method). **Statistically significant models P ≤0.001 (details in Supplementary material).
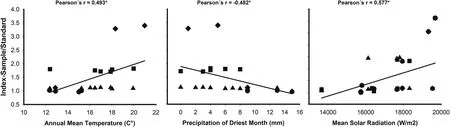
Fig. 3. Pearson correlations between bioclimatic variables (Annual mean temperature (BIO1), Precipitation of the driest month (BIO14), Mean solar radiation) and Relative DNA content ratio(RCR)for four Larrea species.*Statistically significant correlations P ≤0.05.Triangles up:Larrea divaricata,squares:L.cuneifolia,circles:L.nitida,romboids:L.tridentata.
4. Discussion
Our results are consistent with the hypothesis that selective climatic pressures in the genusLarreamight have facilitated repeated whole genome duplication events in North America(Hunziker et al.,1972;Lia et al.,2001;Laport et al.,2013).In South America, reticulate evolution as allopolyploidization by hybridization could have arisen in an environmentally-dependent way that has resulted in the observed correlations between climate and genome size.Studying the degree of relative DNA content changes among the four more abundant species ofLarreathroughout their distributions in North and South American deserts, we found differences in relative DNA content (RCR) and monoploid genome value among species. The small variance observed in their relative DNA content could be potentially helpful to identify species by their genome size. We acknowledge that relative genome size values were calculated based on DAPI staining AT bases. Data on interspecific geographic AT content variation would be necessary to account for this bias.However,the AT bases variation would slightly affect the general patterns observed in the present study.
All studied species showed a correlation between relative DNA content and key temperature and precipitation gradients.L nitidahas the lowest DNA content among the diploid species and also the lowest variation;along with differential morphological characters;enough so that it is classified in another section of the genus(Hunziker and Comas, 2002).L. cuneifolia, as an allotetraploid shows intermediate values of relative DNA content, between hexaploidL. tridentataand the diploid species. The genomic composition ofL. cuneifoliais still unknown, but it has been suggested to have originated from diploid genomes (Lia et al., 2001),whose relative genome sizes should be added together to get comparative theoretic allotetraploid size.On the other extreme,the three populations ofL. tridentatasampled for this study (New Mexico-Chihuahua, Baker-Mojave and Tucson-Sonora), have the highest DNA content, and are inferred to contain diploid and hexaploid cytotypes, respectively. We only sampled 6x individuals in Tucson(Sonora desert),which is not a surprise since Laport et al.(2012) reported several broadly interdigitated populations of tetraploids and hexaploids throughout central Arizona.
Temperature,precipitation and solar radiation have been found to be influential predictors of plant distribution; as higher DNA content tend to occur in harsher climates (Stebbins,1985; Poggio et al., 2014; Rice et al., 2019). Due to the large and disrupt latitudinal span of the genusLarrea(22-46°S in South American Monte Desert,plus scattered sites in Bolivia and Peru,and 20-35°N in North America),environmental selective pressures are expected to differentially affect species inhabiting different portions of the climatic range. Consistent with this hypothesis, inLarrea,climatic variables are correlated with relative DNA content as well.We found that South AmericanLarreaspecies are more likely to have higher DNA content in sites with lower annual precipitation, higher solar radiation,and temperature.L.cuneifolia,the only polyploid species in South America, has the same climatic breadth as the diploidL. divaricata, but they differ in how their RCR relates with climate.The South American allotetraploid shows no correlation between DNA content and environment. Unexpectedly, RCR inL. cuneifoliadecreases with solar radiation,probably as a result of the interaction of this climatic variable and elevation.Meanwhile,significant levels of variation in DNA content are observed along the distribution ofL.divaricatawith latitude and precipitation of the driest month.The correlation between genome size and latitude suggests adaptation to varying climatic conditions at different latitudes, favouring plants possessing certain genome sizes if they are able to tolerate local environmental conditions that might constrain their fitness(Ohri et al., 1986; Bottini et al., 2000; Leong-ˇSkorniˇckova′ et al.,2007; Zhang et al., 2019). The observed pattern might also respond to genetic variation of local population's genomes in combination with DNA amount.Further studies on genetic variation should be conducted over the range of the species to better understand the selection forces acting on the different species.L.nitidaalso partially shares its geographic range with the other South AmericanLarreaspecies, although, it is found only in colder and more humid environments, being a species with high habitat specificity. DNA content inL. nitidaincreases with elevation and decreases with increasing temperature and precipitation. In the North American species(L.tridentata)RCR,along with ploidy level,is positively correlated with temperature and negatively correlated with precipitation (i.e., individuals from drier sites with higher temperatures have larger genomes) in agreement with global patterns reported by Rice et al.(2019).
The reduced set of temperature and precipitation proxies selected here, annual mean temperature, precipitation of the driest month, and solar radiation, differs for South and North AmericanLarreacongeners. In South America, DNA content is negatively correlated with precipitation and positively correlated with temperature.Larreaspecies are hypothesized to have responded to heterogeneous environments through diversification into different species during the Neogene (Lia et al., 2001). Along with this diversification,alloploidization arose by hybridization at least once. The allotetraploid,L. cuneifolia, might have had an advantage at the time of its origin, although today its range overlaps withL.divaricata.The harsh environment of a climatic refugia may have triggered the sympatric diversification via hybridization betweenL. divaricataand other congeneric species that formedL. cuneifolia. In contrast,L. tridentata, is hypothesized to have evolved three ploidal races via autoploidization in the North American deserts during the Quaternary (Lia et al., 2001), potentially as a response to selective pressures due to climatic variation as the species reached North America from South America. This strategy,i.e.whole genome duplication,has been proposed to play a major role in rapid plant evolution and as an important determinant of invasiveness facilitating the establishment in novel environments (Te Beest et al., 2012).
Our results show that both South American and North AmericanLarreaoccupy a wide range of climatic conditions, but there are differences in climatic ranges among species of the genus.We found thatL.nitidawith its small genome size and low DNA content variation,tolerates lower temperatures and low levels of solar radiation,while growing in localities with more precipitation.L.divaricataandL.cuneifoliahave relatively high DNA content variability along their respective distributions and show high tolerance for a range of temperatures and precipitations.L.cuneifolia's climatic envelope is characterized by lower precipitation and higher solar radiation than the other species. The literature suggests that polyploids might tolerate harsher environmental conditions compared to diploid progenitors due to their greater potential fitness (Maceira et al.,1993;Petit and Thompson,1999;Zhang et al.,2019).L.tridentatais exposed tomoresevereclimatic conditions than the South American species; temperatures in North American deserts are higher and precipitation much lower than those found in South America. The climatic envelope forL.tridentatais characterized by high values of temperature and solar radiation along with low precipitation,harsh conditions that might have been selective drivers for polyploidization.L.divaricataandL.tridentataare sister taxa(Lia et al.,2001)andL.divaricatahas a similar monoploid genome value(Cx)and relative nuclear DNA content(RCR)to diploidL.tridentata.This might indicate that the extra DNA inL.tridentatatetraploids and hexaploids is not random,suggesting selection for polyploidization,as postulated forHippeastrum spp.(Poggio et al.,2014).These duplicated genomes can undergo divergent evolution for new functions, giving polyploids ecological benefits that contribute to their success in nature(Soltis and Soltis, 2000; Comai, 2005). Also, as changes in DNA content can influence an organism's ecological amplitude or tolerance,have an important role in gene regulation,or the acquisition of new genetic functions,genome duplication inL.tridentatamay have allowed the species to survive in harsher environments (i.e. drier and hotter). Different species respond in a different fashion to the same climatic conditions.However,differences in DNA content can influence the ecological breadth of the species.L.tridentatahas three cytotypes and grows under much higher temperature than the South American species and in localities with very low precipitation.The ploidal volatility ofL. tridentatamight have provided adaptive advantages in those extreme environments. Energy availability,determined by temperature,light,and water,has been proposed as the primary driver of patterns of species diversity (Rohde, 1992;Hawkins et al., 2003; Willis et al., 2009; Flenley, 2011). Consistent with this hypothesis,temperature has been identified as a correlate of molecular evolutionary rate in plants(Davies et al.,2004),as has water availability(Goldie et al.,2010).So,it is reasonable to expect that plants at drier,hotter and with higher solar radiation environments (i.e. harsher environments) that undergo processes of increasing DNA content may also increase ecological amplitude or acquisition of new genetic functions.
5. Conclusions
In this study we shed light on how DNA content variation in South American desert vegetation taxa played an important role in structuring Western Hemisphere dryland plant communities.Pointing out how South American deserts have been understudied.Correlation between environmental factors and relative DNA content reveal a potentially adaptive role for intraspecific genome size variation. In some cases, repeated whole genome duplication events in plant species might have a selective advantage in harsh environments where climate pressures are high.On the other hand,reticulate evolution as allopolyploidization via hybridization might arise climate-dependently showing particular climate - genome size signals. Increasing DNA content and/or ploidy level, have played a major role in plant evolution, conferring plants a genetically advantageous character,i.e.giving them the ability to adapt to a wider range of climatic conditions.
Author contributions
C.P.S. designed the study and managed the project. M.T. and C.P.S. collected samples. R.V.R. prepared DNA samples and performed cytogenetic analyses along with R.U.and T.U.Data analyses were performed by R.V.R.,C.P.S.and R.U.C.P.S.and R.V.R.wrote the manuscript. M.T. and T.U. modified and improved the manuscript.All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was supported by Argentine National Found for Science and Technology,under Grant PICT 2014-3478- BID and 2019-0149 BID;and Council of Science and Technology-CONICET under Grant PIP-0712. R.V.R., M.T., and C.P.S. are members of CONICET.Grant Agency of the Czech Republic 17-12420S. We thank Peter Smouse and two anonymous reviewers for helpful comments that improved the quality of our manuscript. Thanks also to Thomas Kitzberger, Julieta Betinelli and Daiana Jaume, Carolina Calvi~no,Laura Zalazar, who collaborated in field work, sample processing,and downloading climatic data respectively.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2021.11.006.

- 植物多樣性的其它文章
- GUIDE FOR AUTHORS
- Taxonomic synopsis of Berberis (Berberidaceae) from the northern Hengduan mountains region in China,with descriptions of seven new species
- Plant invasions facilitated by suppression of root nutrient acquisition rather than by disruption of mycorrhizal association in the nativeplant
- SSR markers development and their application in genetic diversity evaluation of garlic (Allium sativum) germplasm
- Mapping the habitat suitability of Ottelia species in Africa
- Adaptive responses drive the success of polyploid yellowcresses(Rorippa, Brassicaceae) in the Hengduan Mountains, a temperate biodiversity hotspot