
Mapping the habitat suitability of Ottelia species in Africa
2022-10-12 06:48:34BonifacNgargaJohnNziJosphatSainaMarwaWasmHalmyJinMingChnZhiZhongLi
植物多樣性 2022年5期
Bonifac K. Ngarga , John M. Nzi , Josphat K. Saina ,Marwa Wasm A. Halmy , Jin-Ming Chn , Zhi-Zhong Li ,*
a Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan, 430074, China
b Center of Conservation Biology, Core Botanical Gardens, Chinese Academy of Sciences, Wuhan, 430074, China
c University of Chinese Academy of Sciences, Beijing,100049, China
d Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, 666303, China
e Department of Environmental Sciences, Faculty of Science, Alexandria University, Alexandria, 21511, Egypt
A B S T R A C T Understanding the influence of environmental covariates on plant distribution is critical, especially for aquatic plant species. Climate change is likely to alter the distribution of aquatic species. However,knowledge of this change on the burden of aquatic macroorganisms is often fraught with difficulty.Ottelia, a model genus for studying the evolution of the aquatic family Hydrocharitaceae, is mainly distributed in slow-flowing creeks, rivers, or lakes throughout pantropical regions in the world. Due to recent rapid climate changes, natural Ottelia populations have declined significantly. By modeling the effects of climate change on the distribution of Ottelia species and assessing the degree of niche similarity, we sought to identify high suitability regions and help formulate conservation strategies. The models use known background points to determine how environmental covariates vary spatially and produce continental maps of the distribution of the Ottelia species in Africa. Additionally, we estimated the possible influences of the optimistic and extreme pessimistic representative concentration pathways scenarios RCP 4.5 and RCP 8.5 for the 2050s. Our results show that the distinct distribution patterns of studied Ottelia species were influenced by topography(elevation)and climate(e.g.,mean temperature of driest quarter,annual precipitation,and precipitation of the driest month).While there is a lack of accord in defining the limiting factors for the distribution of Ottelia species, it is clear that water-temperature conditions have promising effects when kept within optimal ranges. We also note that climate change will impact Ottelia by accelerating fragmentation and habitat loss. The assessment of niche overlap revealed that Ottelia cylindrica and O. verdickii had slightly more similar niches than the other Ottelia species. The present findings identify the need to enhance conservation efforts to safeguard natural Ottelia populations and provide a theoretical basis for the distribution of various Ottelia species in Africa.
Keywords:African freshwater bodies Climate change Ecological niche modeling Habitat suitability Niche overlap
1. Introduction
Climate change is predicted to reduce biodiversity and alter the distribution, productivity and composition of most ecosystems(Hoekstra et al., 2005; Butchart et al., 2010; Hanski 2013; IPCC,2014). Aquatic ecosystems, which are among the most diverse and complex, are especially challenged by climate change and human activities, resulting in habitat loss, degradation, and fragmentation(Murphy et al.,2019).Formulating effective conservation strategies for aquatic ecosystems requires a better understanding of current aquatic biodiversity and species distributions(Bailie 2004;Wilson et al.,2006;McCarthy et al.,2008),and how suitable habitat will change in the future(Pressey et al., 2007;Nzei et al., 2021).
Tropical Africa is characterized by abundant reservoirs that include basins such as Lake Victoria, Congo, and Zambesi Basins,and large rivers such as the Niger,Zambezi, and Congo(Lewis and Berry, 1988). The availability of these vast, interconnected water systems provides an excellent opportunity to assess the effects of climate change on aquatic ecosystems (De Dominicis et al., 2015).Moreover, this region is characterized by a variety of climates,including semiarid, desert, tropical monsoon, and equatorial climates.In addition,with its unique topography,different ecoregions in Africa have different environmental characteristics (Murphy et al., 2019), and it is anticipated that every plant species in the region will show a complex feedback mechanism to climate change(Corlett, 2016). Over the last few decades, Africa has experienced regional and continental climate change, which has significantly impacted the region's water systems (Cav′e et al., 2003; Serdeczny et al., 2017). Future climate projections have suggested that precipitation patterns will change (Serdeczny et al., 2017; Shepard,2019), heat waves will increase, sea levels will rise, and river flow direction will reverse(IPCC,2014).Consequently,by 2050 Africa is predicted to lose ~17% of its drainage (Nyong and Niang-Diop,2006; Misra, 2014). These changes foretell significant potential alterations in the ecology and distribution of wetland species,coupled with expansion or contraction and shifts of ranges where species occur (De Wit and Stankiewicz, 2006). Even so, these wetland species face risks of losing their habitats, and accelerated climate change may drive them to extinction (Corlett, 2016;McLaughlin et al.,2017).
One group of aquatic plants that is affected by both human activities and climate change is the genusOtteliaPers., which includes aquatic species possessing a wide distribution in tropical, subtropical, and temperate regions around the world (Cook et al., 1983; Cook and Urmi-K¨onig, 1984).Otteliaspecies are typically found in shallow lakes, rivers, streams, and water pans due to their preferred inland water habitats (Cook and Urmi-K¨onig, 1984; Li et al., 2020a). These water bodies are frequently exploited for irrigation and polluted by agriculture and the daily routines of locals (Ngarega et al., 2021a), which together may result in habitat degradation, contributing to population declines inOtteliaspecies. While the classification ofOtteliaremained ambiguous until the beginning of the 21st century, the ~23 species that make up the genus are now well documented and have been resolved as a robust monophyletic clade (Li et al.,2020b). In addition, recent cryptic speciation among specific species has heightened interest in further research on the genus(Ito et al., 2019; Li et al., 2019, 2020a, 2020b). Despite the high genetic diversity of theOtteliaspecies(Zhai et al.,2018;Fan et al.,2019; Guo et al., 2019; Li et al., 2019; Ngarega et al., 2021a), few studies have used georeferenced data and statistical models to assess the effects of climate change on the expansion or contraction ofOtteliaspecies’ geographical distributions (Guo et al., 2019). For instance, Guo et al. (2019) used MaxEnt to predict the distribution ofOttelia acuminatain China. The future projection results showed a generally stable habitat under the extreme greenhouse gas emission scenario, with a slight poleward range shift. Africa contains approximately 13 species ofOttelia(Symoens et al., 2009; Li et al., 2020b). However, no studies have yet modeled the distribution of AfricanOtteliaspecies. Therefore assessing the factors that influence the distribution and diversification ofOtteliain Africa may serve as a case study for how freshwater environments respond to climate change.
Ecological niche models(ENMs)are powerful tools for assessing the impacts of climate and environmental change on species, as well as their habitat suitability(Elith and Leathwick,2011;Ngarega et al., 2021b; Wan et al., 2021). Among the presence-only models,the maximum entropy (MaxEnt) is preferred and has been successfully used to forecast the distribution of wetland species(Phillips et al., 2006; Elith et al., 2011; Nzei et al., 2021). Furthermore, MaxEnt has been shown to outperform genetic algorithms and regression models in ENMs(Heikkinen et al.,2006;Elith et al.,2009) and could account for the complex interactions between geographical features that are responsible for the absence or presence of water basins (Phillips et al., 2006). MaxEnt can also predict species’ habitat suitability in unknown wetlands/basins based on presence-only data (e.g., Guo et al., 2019).
In this study, we used MaxEnt to model the current and future geographic distribution ofOtteliain Africa in response to moderate and extreme greenhouse gas emission scenarios (i.e., RCP 4.5 and RCP 8.5 for the period 2050). We aimed to identify the environmental factors that limit the distribution ofOtteliain the region.Due to varied ecological requirements,we hypothesized that theseOtteliaspecies distribution patterns would differ.
2. Materials and methods
2.1. Study area and species
Africa contains approximately 13 species ofOttelia(Symoens et al., 2009; Ngarega et al., 2021a). In the current study, the distribution of seven AfricaOtteliaspecies was modeled, viz.Ottelia cylindrica(T.C.E.Fr.) Dandy, O. muricata(C.H. Wright) Dandy, O.kunenensis(Gurke) Dandy, O.fischeri(Gurke) Dandy, O. ulvifolia(Planch.) Walp, O. exserta(Ridl.) Dandy andO. verdickiiGurke.Modeling the distributions ofO. luapulanaSymoens,O. lisowskiiSymoens, O. scrabaBaker,O. scrabaChiov.,O. brachyphyllaGurke.andO. alismoides(L.) Pers. was not possible because of the limited number of occurrences.For accurate modeling and the production of ftiting models, a species requires more than three occurrence records(van Proosdij et al., 2016).
2.2. Collection of occurrence data
Occurrence and locality data of the AfricanOtteliaspecies were obtained from the Global Biodiversity Information Facility (GBIF,http://www.gbif.org/, accessed on March 2020), TROPICOS (http://www.tropicos.org/, accessed on March 2020), Zambia Flora(https://www.zambiaflora.com/,accessed on March 2020),hitherto published literature (Kennedy et al., 2015) and the authors’ field notes(2018-2019).Notably,there is substantial ambiguity present in the taxonomical literature for the genusOtteliain Africa due to the synonymous use of various names to identify one species (e.g.O.ulvifoliaandO.exserta;see Ferrer-Gallego et al.,2016).Therefore,to avoid any misperception arising from this taxonomic uncertainty, only the literature that listed specific epithet according to Plant List (IPNI; The Plant List, 2010) was considered, and all synonyms that refer to one species were discarded.We cross-checked all herbarium specimens for probable misidentification and noted the locality information on label data. For the herbarium records that lacked geographic coordinates but had locality occurrence descriptions,we used Google Earth?to georeference them at three decimal degrees of accuracy.Once the data was obtained,including a combined data set of 631 occurrence records for all the species,spatial rarefying was performed for the localities of eachOtteliaspecies by using R package “spThin” v.0.1.0 (Aiello et al., 2015) to reduce the autocorrelation between the points matching the resolution of our climatic data, i.e., 2.5~arc min. The number of occurrences that remained and were used for modeling was as follows:22 forO.cylindrica,91 forO.exserta,12 forO.fischeri,24 forO. kunenensis, 57 forO. muricata, 274 forO. ulvifolia, and 33 forO. verdickii(Table S1; Fig.1).
2.3. Environmental variables
Climatic data variables attained from the period 1970-2000 were obtained from the WorldClim v.2.1 database at a resolution of 2.5~arc min from the equator(Fick and Hijmans,2017).This data set(19 bioclimatic variables,Table S2)includes a summary of average,extremes, and seasonality patterns of precipitation and temperature. To ensure variable independence and to limit the impact of multicollinearity and overfitting on the model,Pearson correlations among the 19 bioclimatic variables were calculated based on a threshold-dependent variance inflation factor(VIF)correlation at a threshold of 0.7 using the “vifcor” function in usdm package in R v.3.6.2 (R-Core-Team, 2019). In total, nine bioclimatic variables were selected from the VIF correlation analyses(Table S3).Otteliais an aquatic genus,and due to its dependency on water,we included precipitation of the driest quarter(Bio17)and annual precipitation(Bio12) as relevant to water availability for the genus. We also incorporated elevation (http://srtm.csi.cgiar.org/) as an extra variable becauseOtteliais affected by geography (Symoens, 2009); in addition, we extracted slope using the same variable with the Surface Analyst Tool in ArcGIS v.10.5.The thirteen selected variables were used for the subsequent analyses(Table S2).For global climate models (GCMs), we utilized the Community Climate System Model4(CCSM4;Gent et al.,2011),which is considered suitable for the African region (McSweeney et al., 2015) for the period 2041-2060, commonly referred to as period “2050”. Two RCPs were selected for the CCSM4 period to represent the moderate(RCP 4.5) and extreme climatic changes (RCP 8.5). RCP 8.5 assumes no climate policies will be undertaken either in the future or present,and it reflects very high emissions scenarios (~1370-ppm CO2equivalent by 2100; van Vuuren et al., 2011). All future climatic layers used the spatial resolution of 2.5~arc min and were retrieved from WorldClim v.1.4 (Hijmans et al., 2005). Due to future complexities and obtaining data for these periods, the topographic variables(elevation and slope)remained kept constant for current and future projections.
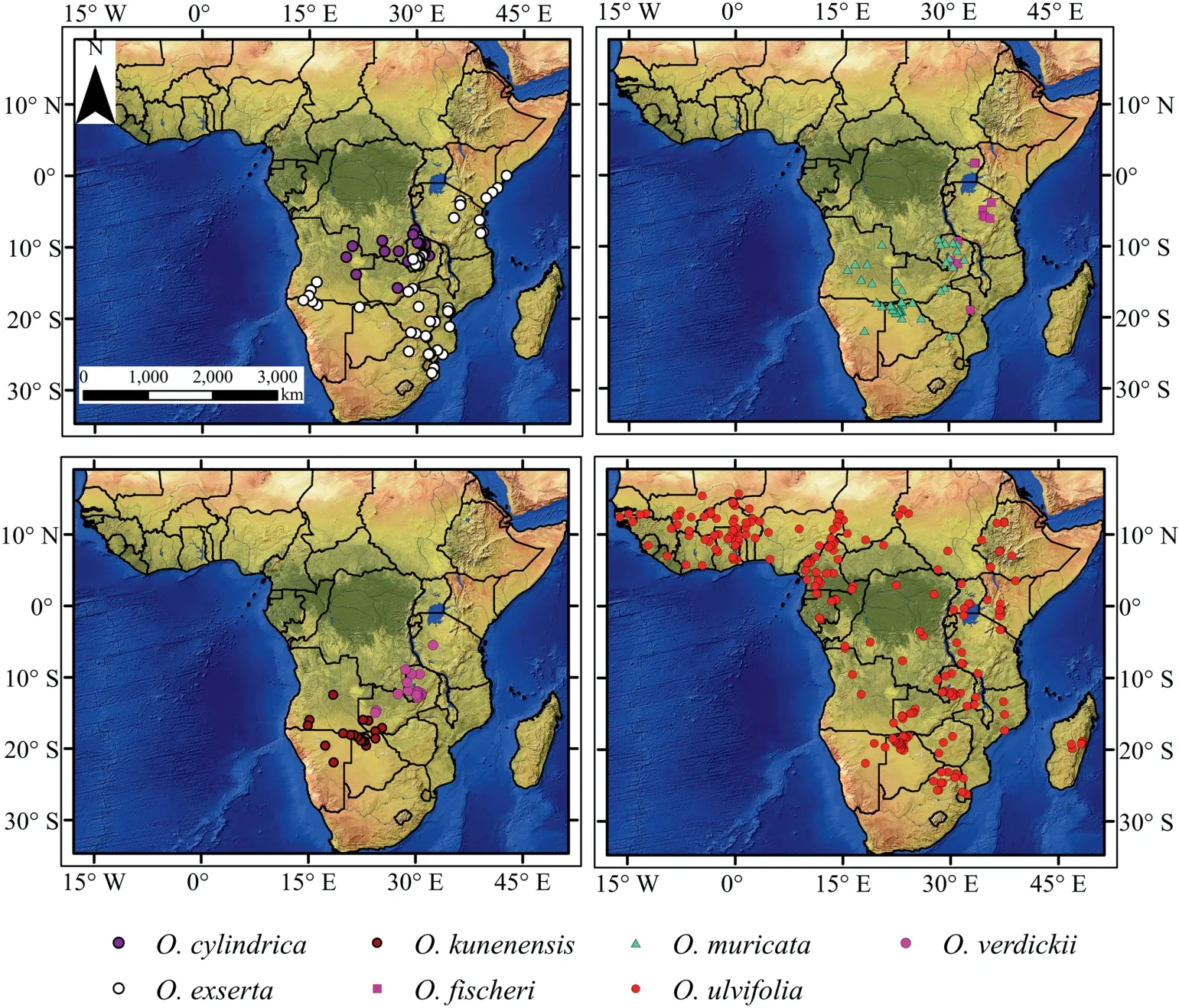
Fig.1. Map of the occurrences of Ottelia spp.Occurrences of Ottelia cylindrica(n=22),O.exserta(n=91),O.fischeri(n=12),O.kunenensis(n=24),O.muricata(n=57),O.ulvifolia(n = 274), O. verdickii (n = 33) used for model development.
2.4. Ecological modeling and validation
The maximum entropy algorithm applied in MaxEnt v.3.4.3.e(Phillips et al.,2006)was used to model the distribution likelihood of sevenOtteliaspecies in response to various environmental constraints (Table 1). We used the ENMeval package in R v.3.6.2(Muscarella et al., 2014) to perform a tuning experiment of our MaxEnt models. Using defaults for MaxEnt has resulted in poorperforming models (Radosavljevic and Anderson, 2014; Warren and Seifert, 2011). The ‘block method’, which divides data (presence points) into k = folds (Wenger and Olden, 2012), was employed to make the locality data into independent spatial test and training data sets.The regularization multipliers were set from 0.5 to 5 with increments of 0.5 in the k of MaxEnt models. The feature combinations used were L, H, LQ, LQH, LQHP, and LQHPT;where P = product, H = hinge, T = threshold, Q = quadratic, and L =linear.The tuning experiment was applied for all sevenOtteliaspecies, and then MaxEnt was run with the following parameters:50 bootstrap replicates and 5000 iterations. The number of background points influences model predictions;therefore,it is advised that a high number of points,generally 10,000 or more,be used to reflect the background environments of species being contrasted with presences depending on the geographical scale of the ecological questions of interest(Phillips et al.,2008;Barbet-Massin et al., 2012). Thus, for building theOtteliadistribution models, we used MaxEnt default 10,000 background points as recommended by Phillips et al.(2008)and Barbet-Massin et al.(2012).Validation of the models included performing the receiver operating characteristics(ROC)curve analysis to estimate the values of AUCtest and AUCtrain (Warren and Seifert, 2011) and the True skills statistics(TSS)(Allouche et al.,2006).We also included the jackknife tests to analyze the relative importance of the selected variables. The model results were given as averages of the fifty replicates and converted into binary layers with values of 1 (presence) or 0 (absence) for each species, using the maximum training and sensitivity plus specificity (MTSS) as a threshold. MTSS is recommended as a conservative approach that minimizes commission and omission errors (Liu et al., 2016; Guisan et al., 2017). This analysis was performed in ArcGIS v.10.5. Thus, one baseline (current) and two potential distribution layers were created for each species based on the two scenarios and one GCM. Future habitat suitabilities were based on the current records and variables described before.
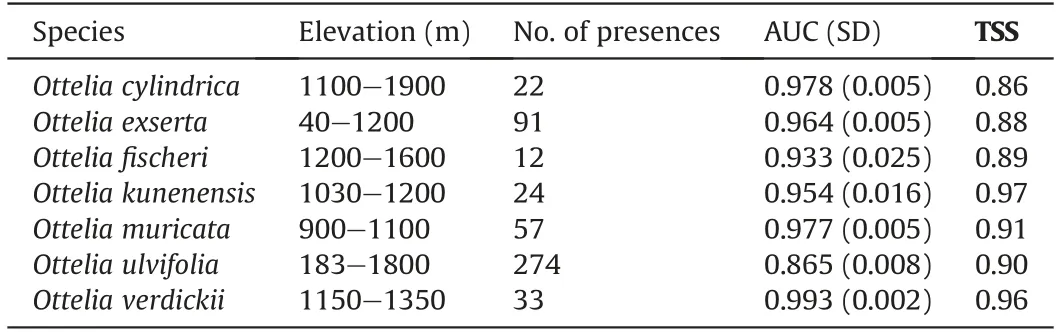
Table 1 Characteristics of the Ottelia species’ spatial records were used as predictors to model the tropical African freshwater SDMs,including model performance based on mean AUC and TSS values. Standard deviation in parentheses.
2.5. Distributional changes within binary models
To assess the influence of climate on the distribution ofOttelia,we analyzed the distributional changes in habitat suitability for each taxon between the current and future thresholded binary models.Using SDM toolbox v.2.3(Brown,2014)in ArcGIS v.10.5,the range changes were analyzed with respect to the current period and classified as stable, loss, gain, and unsuitable.
2.6. Niche overlap assessment
The evaluation of niche overlap allows estimating the niche shared byOtteliaspecies. We used Schoener'sD(Warren et al.,2008) statistics to assess the niche overlap of focal species and identity, implemented in ENM tools v.1.4.4 (Warren et al., 2010).These two metrics compare niche overlap and identity of the two species with values from 0 to 1, with 1 signifying resemblance in the two niches and 0,no overlaps(Waren,2008).We excluded 5%of the occurrence data to account for probable errors in the data.Additionally, we evaluated the null hypothesis that the ecological niche models of each species pair were equivalent. The logistic output maps for the sevenOtteliaspecies pairwise pairings were contrasted to identify the degree of geographical overlap between the species distributions.
3. Results
3.1. Model performance and important environmental variables
The predicted model outputs showed high AUC and TSS values(>0.865 and >0.86, respectively), indicating high predictive performance (Table 1).Under the MaxEnt models,the contribution of variables differed among the species (Table 2). For example, Bio12 contributed greatly to the habitat suitability ofOttelia cylindrica,O. ulvifolia, andO. verdickii. Bio9 contributed greatly to the habitat suitability ofO. exserta, O. muricata,andO. kunenensis. Elevation mostly limited the distribution ofO.exserta,O.fischeri,O.cylindrica,andO.verdickii.Bio8,Bio15,and slope contributed least in allOtteliaspecies distribution models. The results of Jackknife analyses showed that when used in isolation,elevation,Bio8 and Bio12 were the most important variables in the distribution models forO.fischeri,O.cylindrica,O.exserta,andO.verdickii(Fig.S1).Jackknife results forO.muricataandO.kunenensismodels revealed that Bio8 and Bio12 were significant contributors to the models. For theO.ulvifoliamodel,Bio12,Bio13 and Bio9 were identified as the most important factors by jackknife results(Fig. S1).
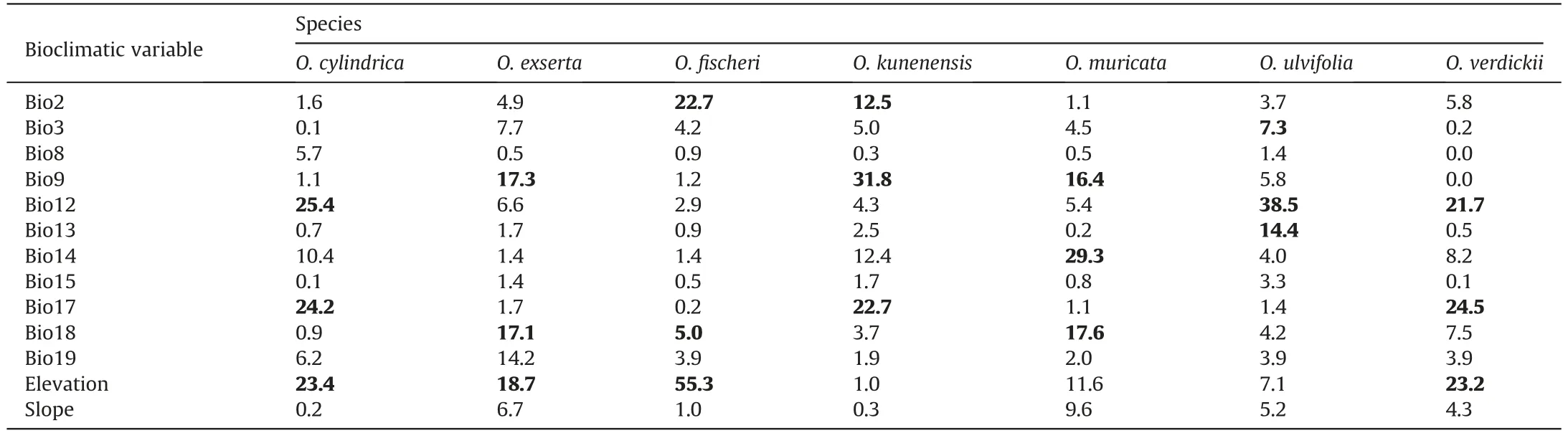
Table 2 Mean relative contribution of each variable to the final model of the current distribution for each Ottelia species studied.
3.2. Predictions of potential distribution under current and future climate change
Our models indicate that current suitable areas for eachOtteliaspecies differ (Fig. 2). Of the seven species examined, the current potential distribution ofO. verdickiiwas the smallest, with highly favorable habitats located in Zambia and western Tanzania.Current potential suitable habitats forO. kunenensisandO. muricatawere similar with highly suitable regions located in the Okavango delta.Moreover, highly suitable habitat forO.fischeriwas identified in large parts of Uganda, Tanzania and northern Zambia.O. ulvifoliawas predicted to have the greatest current potential distribution in Tropical Africa (Fig. 2). In Madagascar, where onlyO. ulvifoliaoccurs, our model revealed suitable high-quality habitats forO. exsertaandO.fischeri, and low-quality habitats forO. cylindrica.Furthermore, the most common region predicted to contain a highly suitable habitat forOtteliaspecies in Africa was the southern hemisphere, including Zambia, Botswana, Angola, and southern Tanzania.
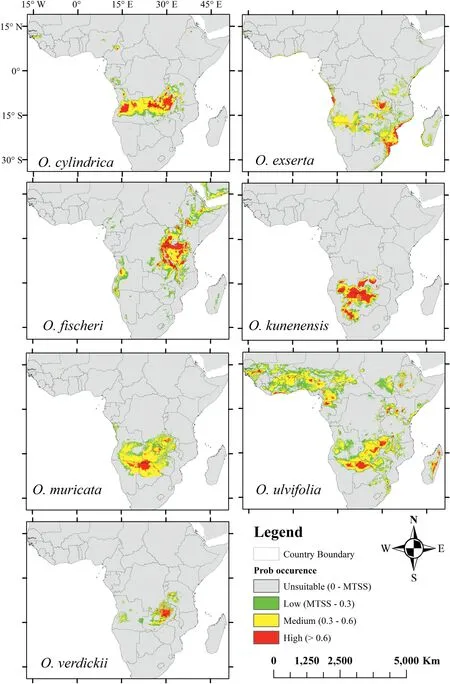
Fig. 2. Current potential distribution of Ottelia species in Africa.
When we modeled potentialOtteliadistributions in response to climate change, the distributions for each species differed under moderate and extreme greenhouse gas emission scenarios(Figs.3 and 4,respectively).By the 2050s,moderate and extreme emission scenarios were predicted to decrease in habitat suitability forOttelia(Figs.5 and 6,respectively).However,an extreme emission scenario was predicted to cause a remarkable loss in habitat ranges (Table S4; Fig. 6). Additionally, under extreme emission scenarios, potentially suitable habitats forO. ulvifoliaandO. exsertain Madagascar and Angola were predicted to be lost(Fig. 6); however, under the moderate emission scenario, these habitats were minimally reduced(Fig.5).Interestingly,O.verdickiishowed the most gain in suitable areas under the extreme scenario (Fig. 6). In both emissions scenarios, we observed that all species ranges are predicted to shift north, as indicated by range expansion (Figs. 5 and 6).
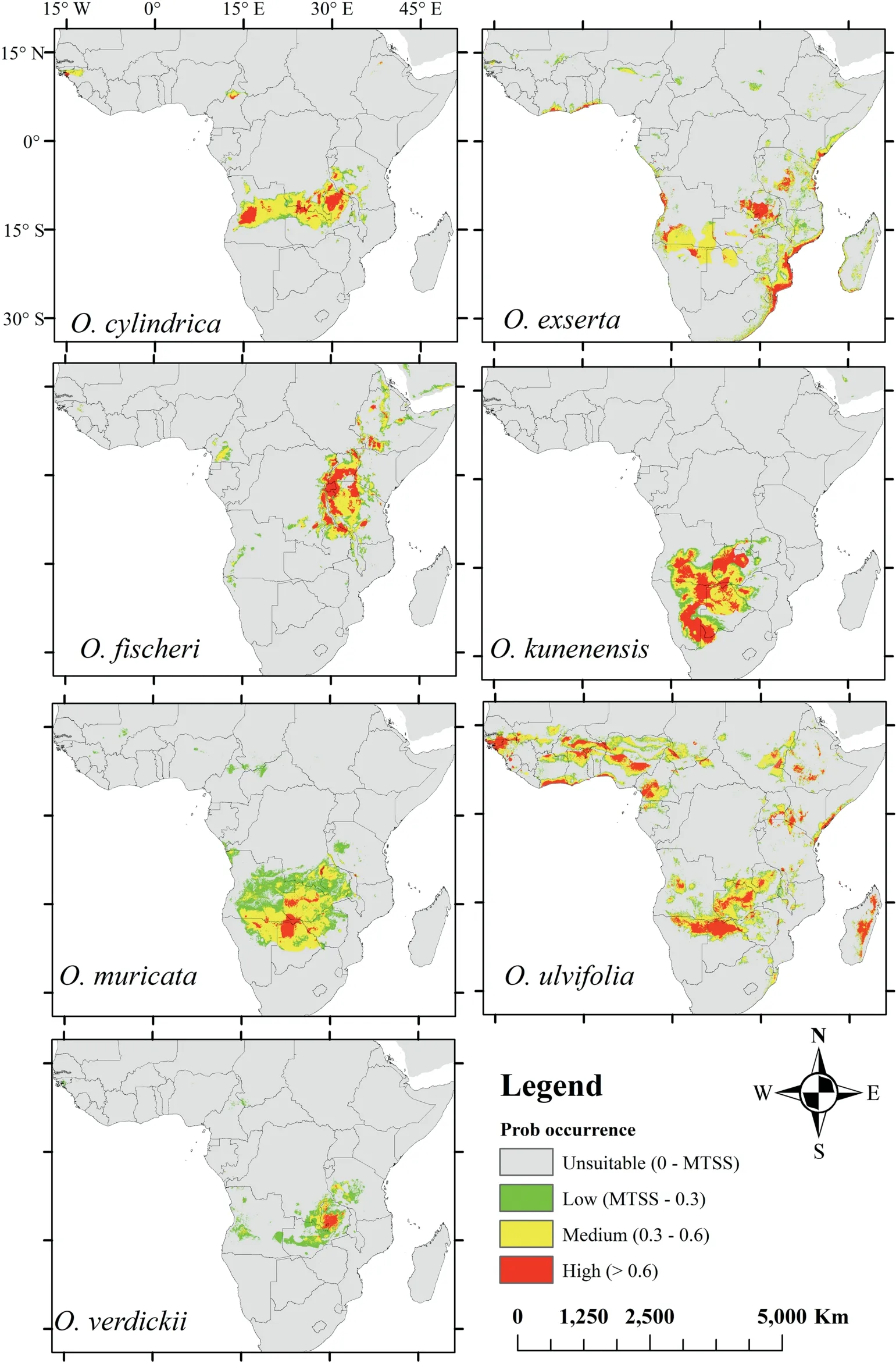
Fig. 3. Predicted potential distribution of the Ottelia species in the 2050s for RCP 4.5 using the MaxEnt models.
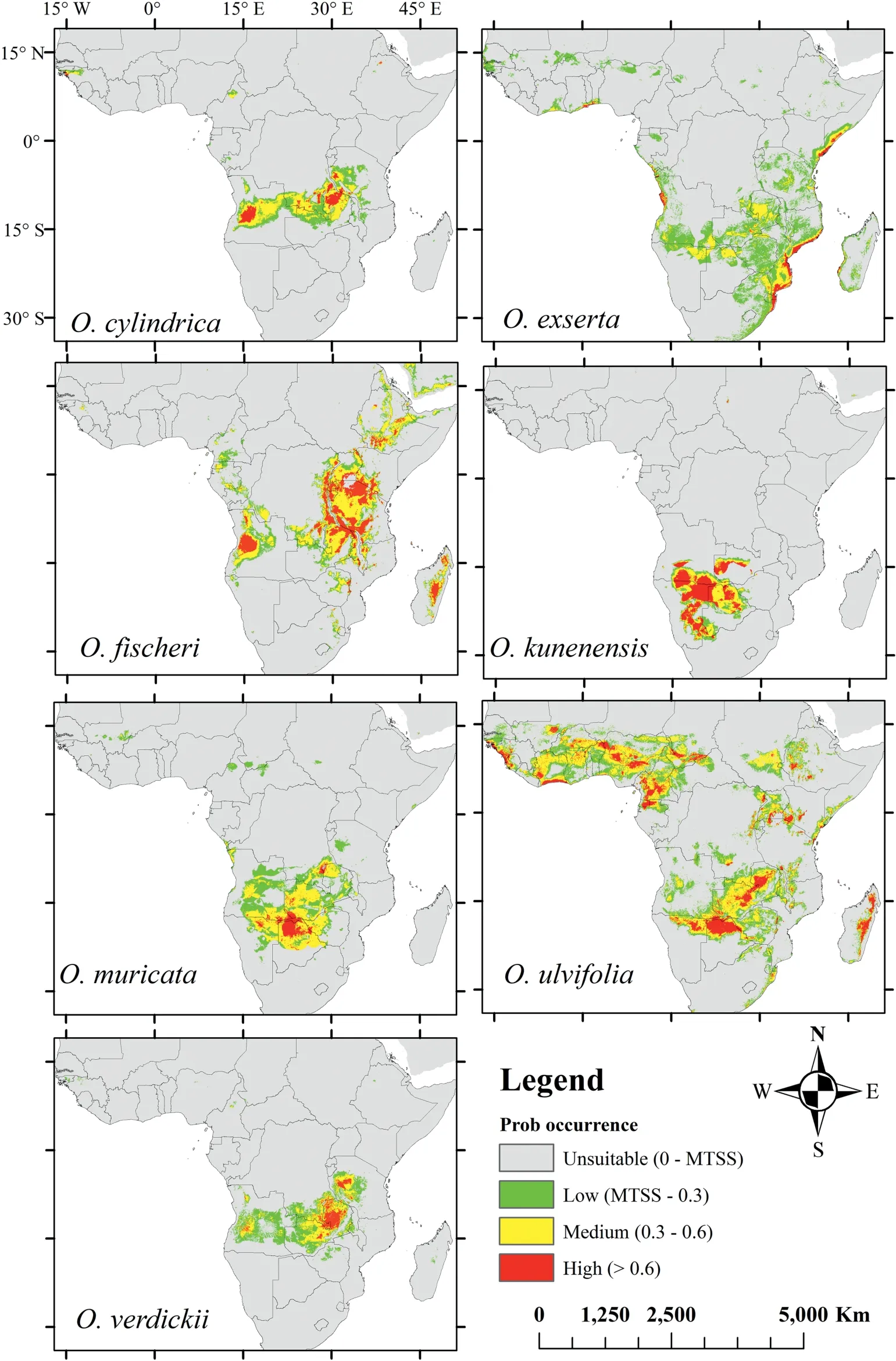
Fig. 4. Predicted potential distribution of the Ottelia species in the 2050s for RCP 8.5 using the MaxEnt models.
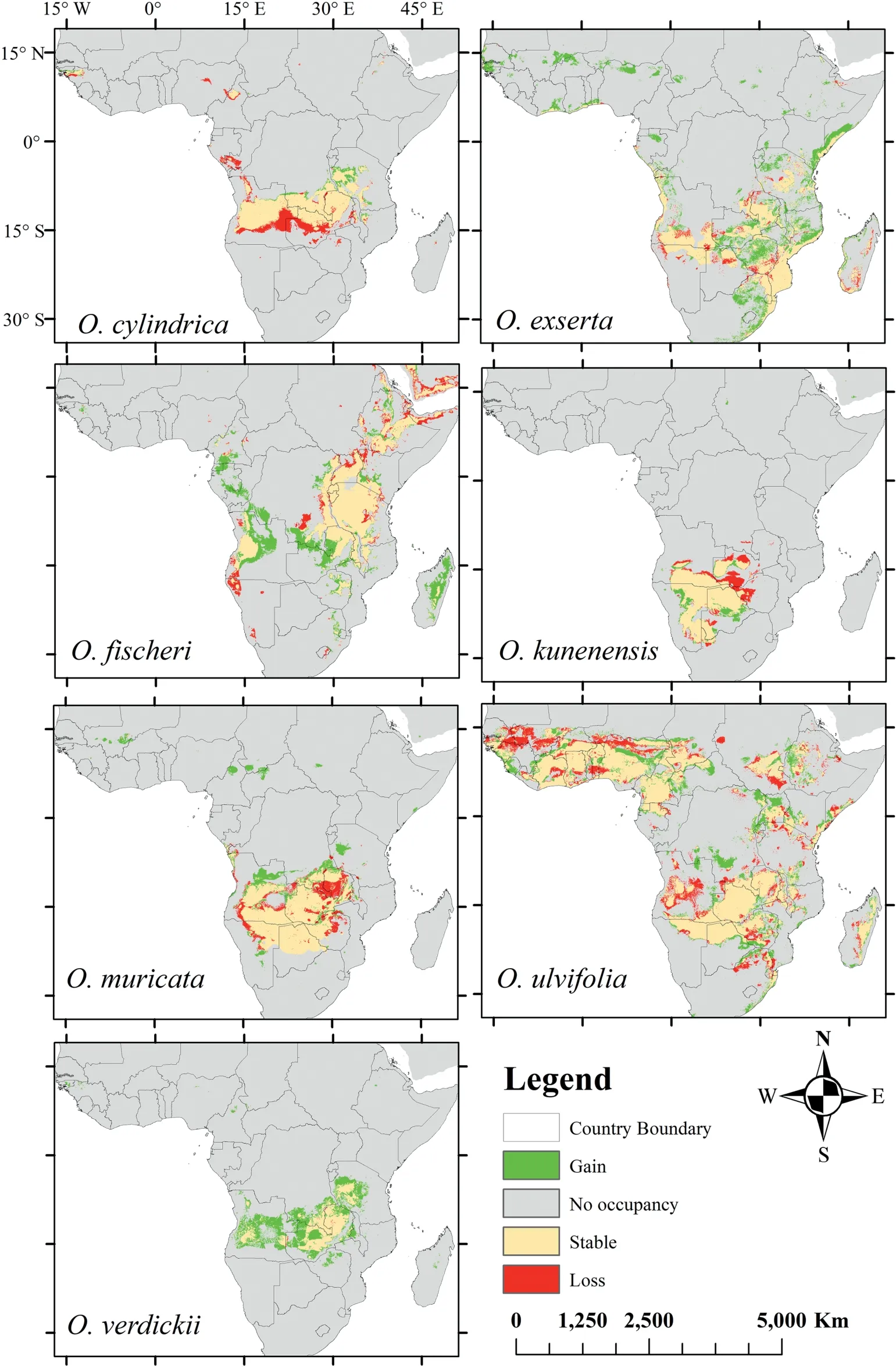
Fig. 5. Predicted change in the distribution of Ottelia species under RCP 4.5 scenario for the 2050s.
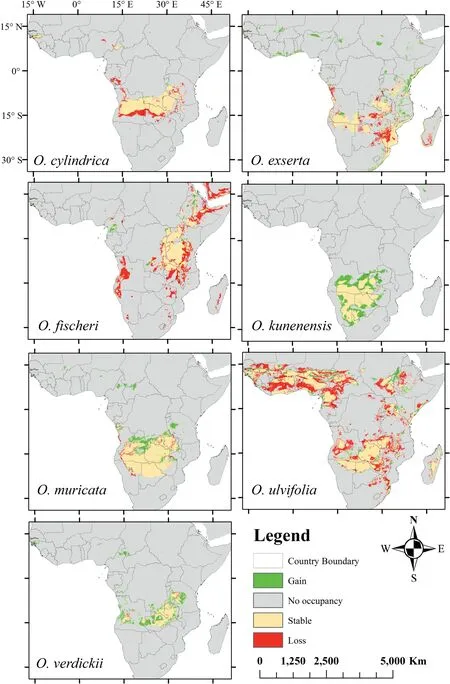
Fig. 6. Predicted change in the distribution of Ottelia species under RCP 8.5 scenario for the 2050s.
3.3. Niche comparisons
According toDstatistics,Otteliaspecies niches overlap considerably(Table 3).O.cylindricaandO.verdickiihad the highest niche overlap (0.651). However, other species, such asO. kunenensisandO.fischeri, occupy different environmental niches. The null hypothesis that assumed that the habitats were equal for pairwiseOtteliaspecies comparisons was not rejected by niche identity tests(Table 3).
4. Discussion
Mapping habitat suitability with an ENM approach is advantageous because it offers critical notes on species management and conservation(Elith et al.,2011).Among the various ENM techniques,MaxEnt has been widely used in aquatic species distributional modeling studies (e.g., Guo et al., 2019; Heneidy et al., 2019; Nzei et al., 2021). To our knowledge, this study is the first to use Max-Ent to predict habitat suitability forOtteliain Africa. Our MaxEnt distribution models had good prediction accuracy (AUC and TSS values >0.86), indicating that environmental variables were key predictors ofOtteliaspecies distribution in Africa(Table 1).
Although the models in the present study displayed excellent overall accuracy, several uncertainties might impact our predictions, including background data (pseudo-absence), sample size and bias(species occurrence localities),and the extent of the study area(Muscarella et al.,2014).For instance,increased sample size enhances prediction accuracy;however,this occurs only up to a limit; then diminishes when an excessively high number of samples is employed (Feeley and Silman, 2011; Muscarella et al.,2014).
Environmental variables directly or indirectly affect ENMs(Austin,2002).In the analyses for the current period,we observed that both the precipitation and temperature variables and elevation limited the distribution ofOttelia. Precipitation and temperature might influence the survival of organisms and their physiology directly.On the other hand,elevation may indirectly affect species abundance and geographical distribution; however, it does not significantly influence their physiology (Duclos et al., 2019). The environmental variables making the highest contribution to the current habitat suitability and distribution ofOtteliadiffered between the species (Table 2). The environmental variables with the largest contributions to most distribution models ofOtteliawere mean temperature of the driest quarter(Bio9),annual precipitation(Bio12), precipitation of the driest quarter (Bio17), and elevation,indicating that these four variables play vital roles inOtteliadistribution. Mean temperature of the wettest quarter (Bio8), precipitation seasonality (Bio15), precipitation of the coldest quarter(Bio19),and slope had little influence on the distribution models forOtteliaspecies(Table 2).Most of Africa receives less than 1000 mm of rain each year on average (Nicholson, 2000). Notably, rainfall decreases with distance from the equator and is negligible in the Sahara (approximately 16°N), eastern Somalia, and southwestern Africa in Namibia and South Africa. On the other hand, rainfall is most plentiful along Madagascar's eastern coast, in sections of eastern Africa's highlands, in vast regions of the Congo Basin and central Africa,along with parts of western Africa's coast,including Liberia, Sierra Leone, and Guinea. This explains the tolerance ofO.verdickii,O.kunenensis,O.cylindrica,O.exserta,andO.ulvifoliato annual precipitation(Bio12;Fig.S1).Previous studies indicate that precipitation and temperature greatly impact the distribution of aquatic species in tropical Africa (Nzei et al., 2021), which is characterized by a variety of climates.This is in line with our simulation results, as regions of suitableOtteliahabitat align with areas that have suitable climates for the persistence ofOttelia(Fig. 2). In contrast, the drier regions (e.g., far north and south poles) are predicted to be unsuitable forOtteliaspecies(Fig.2).These factors and the others included in the current study are possibly linked to ecosystem competence and, particularly, the availability of nutrients (Crossley et al., 2002; Zhang et al., 2020). Additionally, the volume of water in ponds, streams, and lakes or wetlands is also directly connected to precipitation (wet conditions). Climatic variations have been proven to aid evolutionary divergence (Rissler and Apodaca, 2007) by fostering species adaptation to new climatic environs (Iannella et al., 2019). Also, the cosmopolitan distribution ofO. ulvifoliain Africa, supported by our simulations,shows that the species can persist in a broad range of environmental conditions.
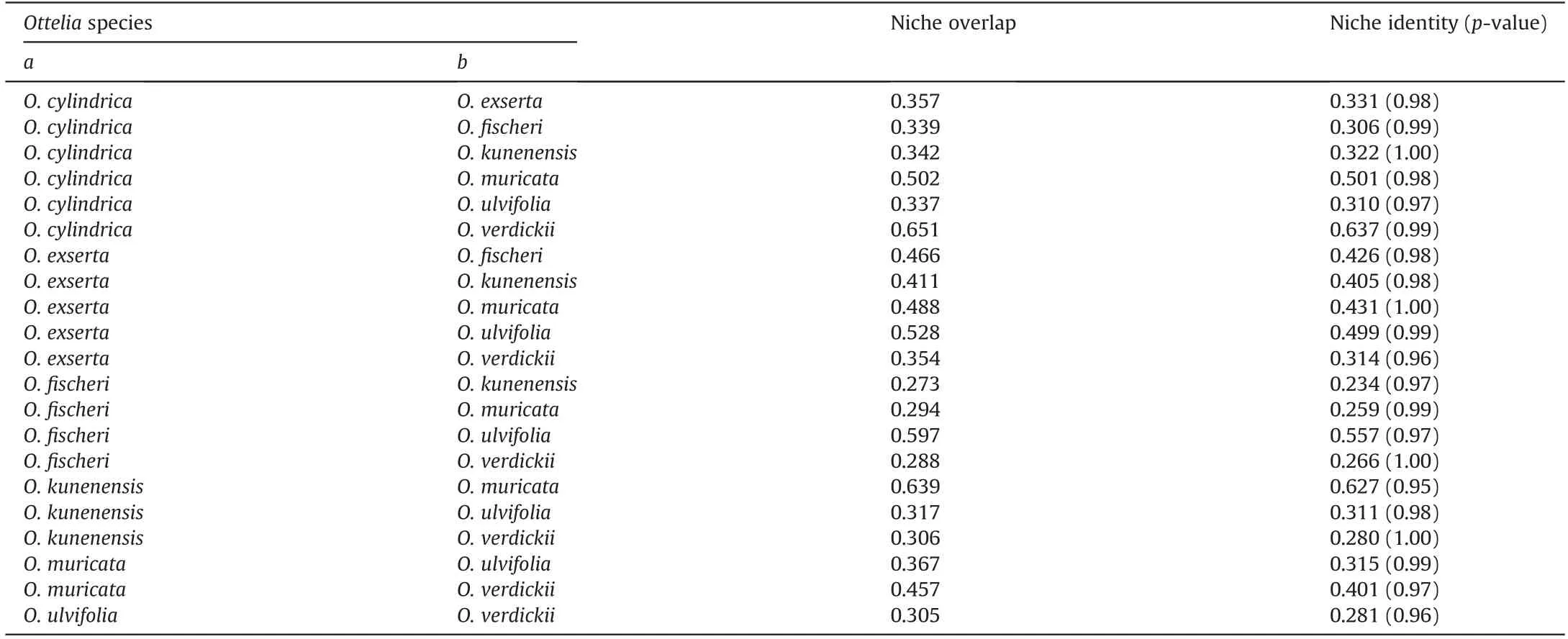
Table 3 Comparisons of ecological niches for Ottelia species. Niche overlap and niche identity tests of species a against species b.
MaxEnt modeling indicated that models of current suitable habitat for the seven AfricanOtteliaspecies are accurate (Fig. 2).Specifically, models indicated that highly suitable habitats are currently located in southern Africa for all species except forO. ulvifolia, which is widely distributed in tropical Africa, andO.fishcheri,which is distributed in central-east Africa.We noted that the range of suitable habitat predicted forO.ulvifoliais larger than that predicted for the otherOtteliaspecies (Table S4), withO. verdickiioccupying the smallest ranges (5,300,853 and 633,504 km2, respectively).In addition,our models indicate that a highly suitable habitat is currentlyavailable in northern and western Africa for threeOtteliaspecies (i.e.,O.fishcheri, O. cylindrica,andO.exserta).These species have not previously been recorded in these regions,indicating that these species may persist in those regions.However,because of the relatively poor ability of aquatic species to disperse, this has not yet occurred (Li et al., 2020a; Ngarega et al.,2021a). Our model likely indicates that southern Africa has the most suitable habitat forOtteliaspecies because the region has appropriate temperatures and an abundance of highly interconnected river systems(Li et al.,2020a;Ngarega et al.,2021a).
Future projections indicate that the response ofOtteliato climate change may be highly variable (Figs. 3 and 4). Although certain species are expected to be largely unaffected by climate change(e.g.,O.verdickii,in RCP 8.5,Fig.6),others,especially in the extreme emissions scenario (RCP 8.5), are expected to lose a significant amount of their appropriate habitat range(Fig.6).The loss of appropriate ranges for a species means,at the very least,a rise in environmental pressure supporting mortality over the establishment in certain areas,which could result in local extinctions(Walck et al., 2011; Parmesan et al., 2015). The different responses to climate change among closely relatedOtteliaspecies also indicate that macrophyte populations may disaggregate in the future,affecting the function and structure of the macrophyte assemblages they formed (Murphy et al., 2019; Alahuhta et al., 2020; García et al., 2020).
The distribution ofOtteliaspecies shows some overlap along their range; thus, someOtteliaspecies have similar niches. Unsurprisingly, our model predicts a continuum ofOtteliadistributions.Given that annual precipitation (Bio12), precipitation of the driest quarter (Bio17), and elevation were significant determinants of habitat suitability for bothO. cylindricaandO. verdickii, it is not surprising that these two species had the most comparable niche overlap and identity (Tables 2 and 3). On the other hand,O. muricata, likeO. kunenensis, is found in productive areas and wetlands in southern Africa(Fig.2).Indeed,our model forecast that the highly suitable niches of these two species overlap in much of the basins around the Okavango Delta and the regions bordering Zambia, Zimbabwe, and Botswana, except for the highly suitable areas in Angola and Namibia (Fig. 2).
Overall, given the wide range of climatic conditions in whichOtteliaspecies are found, it is perhaps not unexpected that niche overlap and identity among these species is predicted to be highly variable(i.e.,from low to moderate,Table 3).Niche separation has been postulated as one of the critical causes that differentiate the distribution of closely related species, mainly through resource competition (Wiens and Graham 2005). In addition, taxa with similar distributional ranges generally utilize habitats differently to prevent complete overlap, reduce competition, and enable coexistence (Schoener, 1974). The theory of competitive exclusion argues that organisms never coexist if their requirements are the same(Hardin,1960).Therefore,ecological/habitat segregation may eliminate interspecific competition, allowing species with comparable requirements to coexist, resulting in a highly diversified population (Schoener,1974).
The present study did not include the effects of pH and nutrient availability of water bodies where species occur or their influence on aquatic species. Therefore, future studies should assess the effects of pH and nutrient availability on the distribution of aquatic macrophytes. Regardless, the observed areas of high habitat suitability for theOtteliaspecies could be observed as potential future refuges.To anticipate the intricate impacts that climate change will have on community assemblages, it is crucial to locate the areas that will most likely persist under adverse future climate change(Monsarrat et al., 2019; Pennino et al., 2020).
Our current study assessed the probable effects of future climate change on the distribution of sevenOtteliaspecies in African freshwater basins. Studies such as this, which use predictive models based on recorded data,established biological parameters,and the best possible climate simulations representing the full spectrum of outcomes, offer policymakers and experts critical knowledge to set goals for mitigation and conservation in the future. The present findings highlight that among the most vital variables that influence the distribution ofOtteliaare elevation,annual precipitation(Bio12),and precipitation of the driest month(Bio14). Projections of future habitat suitability showed that the currently suitable area of theOtteliaspecies would decline by the 2050s under both mild (RCP 4.5) and extreme (RCP 8.5) climate scenarios, with the pessimistic scenario resulting in a tremendous loss.Extensive reductions in suitable habitat in response to climate change will likely threaten or endanger species if management and conservation ofOtteliais not observed. Although currentlyOtteliaspecies are distributed in habitats with satisfactory characteristics,more surveys are encouraged throughout tropical Africa to assess theOtteliaspecies distribution. Although we have successfully modeled the distribution ofOtteliain Africa under a climate change context and showed that the distribution of theseOtteliaspecies would significantly be affected by climate change in the near future,we also note that the species are of “Least Concern” status under the IUCN classification.However,species in this category that have small distributions are usually at high risk of extinction; furthermore,ENM predictions become less reliable for species with small distributions (Schwartz et al., 2006). Therefore species with restricted distributions, for instance,O. verdickii, should be closely monitored.Water is an essential prerequisite forOttelia.Given that the need for its reproduction and dissemination of seeds or propagules are primarily dependent on water, precipitation availability is likely critical forOtteliahabitats.
5. Conclusion
We have demonstrated that global climate change would constitute a severe threat toOtteliaspecies.This study is the first to use MaxEnt to forecast the distribution ofOtteliabased on environmental variables in the African region.According to the present results,the most suitable areas ofOtteliaare located in the eastern and southern African regions. While the current study focused on genusOttelia,particularly the African species,we acknowledge that many aquatic macrophytes need careful consideration. Although we recommend more proactive attempts to safeguardOtteliafrom increasing threats here, we propose that initiatives be made to evaluate and utilize accessible data continuously, together with countrywide level assessments (e.g., Kennedy et al., 2015) to improve the conservation protection of aquatic macrophytes in Africa. Such discussions are currently underway in the African region and are imperative to building preparedness capacity for the ongoing effects of climate change on African waterbodies. Therefore,results from this study are especially pertinent to elaborate the conservation schemes for aquatic macrophytes.Lastly,our findings call for further research into the genusOtteliafrom a variety of viewpoints,including not only the ENMs and ecological aspects but also the genus’ taxonomy, physiology, and molecular barcoding.
Author contributions
B.K.N.,J.M.N.,J.M.C.,and L.Z.Z.conceived the idea and designed the simulations; B.K.N., J.M.C., and L.Z.Z. collected the data; B.K.N.,J.K.S.,and M.W.A.H.presented the methods and analyzed the data.J.M.C. provided the funds for the study. All authors discussed the results and reviewed the manuscript.
Declaration of competing interest
The authors declare no competing conflicts of interest.
Acknowledgments
We are grateful to John M. Ndung’u for early consultations on this study’s statistical approach.In addition,we would like to thank Drs.Valerie F.Masocha(Xishuangbanna Tropical Botanical Garden)and Yeshitila Mekbib(Ethiopian Biodiversity Institute)for thoughtprovoking discussions regarding the study. Thanks are also due to the Kasanka trust for providing logistics during our fieldwork in Zambia. We acknowledge the Department of National Parks and Wildlife,Zambia,for the competent authority of permits during our fieldwork in Zambia.We thank the anonymous reviewers for their suggestions and comments on an earlier draft of our manuscript.This research was fundedfunded by the Strategic Priority Research Program of the Chinese Academy of Sciences(No.XDB 31000000),the National Natural Science Foundation of China (Nos. 32070253 and 32100186), and the Sino-Africa Joint Research Center (No.SAJC201322).
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2021.12.006.

- 植物多樣性的其它文章
- GUIDE FOR AUTHORS
- Taxonomic synopsis of Berberis (Berberidaceae) from the northern Hengduan mountains region in China,with descriptions of seven new species
- Plant invasions facilitated by suppression of root nutrient acquisition rather than by disruption of mycorrhizal association in the nativeplant
- Evolutionary importance of the relationship between cytogeography and climate: New insights on creosote bushes from North and South America
- SSR markers development and their application in genetic diversity evaluation of garlic (Allium sativum) germplasm
- Adaptive responses drive the success of polyploid yellowcresses(Rorippa, Brassicaceae) in the Hengduan Mountains, a temperate biodiversity hotspot