
Transcriptome analysis shows nicotinamide seed treatment alters expression of genes involved in defense and epigenetic processes in roots of seedlings of Picea abies
2022-09-08 06:16:42CeciliaLaurellTorkelBerglundAnnaOhlsson
Journal of Forestry Research 2022年4期
Cecilia Laurell ·Torkel Berglund ·Anna B.Ohlsson
Abstract Nicotinamide treatment of plants and plant cell cultures has been shown to promote defense and decrease levels of DNA methylation.In the present study, we used RNA-seq technology to study overall changes in gene expression induced in roots of 3-month-old spruce ( Picea abies) seedlings grown from nicotinamide-treated seeds to examine the molecular mechanisms underlying the defense promotion.Approximately 350 genes were identified as differentially expressed in roots after the seed treatment.Stress response genes, including transcription factors MYB77 and LHY and two chitinase enzymes, were generally upregulated, whereas genes thought to be involved in epigenetic regulation such as DDM1, known to promote DNA methylation, were present at high frequency among the downregulated genes.Across all samples, the expression of downregulated epigenetic-related genes was highly correlated with the nicotinamide treatment, indicating a common regulation.Our results support an earlier hypothesis regarding a potential role of nicotinamide as a defense-signal mediator.
Keywords DNA methylation·Large-scale gene expression analysis·Nicotinamide·Picea abies · Seed treatment
Abbreviations

ABA Abscisic acid DEG Differentially expressed genes ET Ethylene GO Gene ontology JA Jasmonic acid NIC Nicotinamide PARP Poly-ADP(ribose) polymerase qRT-PCR Quantitative reverse transcription PCR ROS Reactive oxygen species SA Salicylic acid SAR Systemic acquired resistance TE Transposable elements
Introduction
Environmental stresses limit productivity in agriculture and forestry.Plants must adapt to biotic and abiotic stresses to survive.Norway spruce (Picea abiesL.Karst.) is a significant element of European ecology and the forestry industry.Newly planted seedlings often suffer from attacks by pests such as pine weevils.A considerable proportion of the pesticides used in forestry are intended to reduce insect attacks so that newly planted young seedlings survive, but many of these chemical agents have broad ecotoxicity.An alternative approach is to activate and strengthen the plants’ own innate defense systems.
Systemic acquired resistance (SAR) refers to the induction of a strengthened defense response in the whole plant(Luna et al.2012; Conrath et al.2015).Both priming and long-distance hormonal signaling are parts of SAR.Priming is a procedure where plants are induced to become more stress-tolerant via early exposure to a stressor or a defensepromoting signal molecule.Subsequent re-exposures lead to stress responses that are far stronger than the first response(Conrath et al.2015).The plants can acquire a “memory”that protects them not only from the specific stress they have been exposed to, but also to other stresses.UV-C exposure of lettuce, for instance, results in a more salt-tolerant plant(Ouhibi et al.2014).An advantage of the priming technique is that it can be applied to seeds (Jisha et al.2013; Worrall et al.2012).
Stress memory in plants lasts only for a short time period when it is mediated via proteins, RNAs and metabolites.However, stress-induced epigenetic changes, which are considered to be one result of seed priming, can be maintained for longer durations and may even be inherited trans-generationally (He and Li 2018).Epigenetic regulation in plants involves methylation of cytosine in the sequence contexts CG, CHG and CHH in the DNA, as well as histone modifications and other chromatin rearrangements (Bartels et al.2018).It has been demonstrated that the stress-induced DNA methylome is correlated with simultaneous differential gene expression (Dowen et al.2012; Zogli and Libault 2017).Establishment of an epigenetic memory in spruce has previously been discussed around the influence of temperature during embryogenesis (Carneros et al.2017).Other important factors influencing the epitype include the geography and climate of origin, as demonstrated for transcript patterns and DNA methylation in plants ofPopulus(Raj et al.2011)andArabidopsis thaliana(Kawakatsu et al.2016) from different locations.Environmental factors in the habitat of ancestral plants might also pre-adapt some plants to biotic and abiotic stresses via inherited epigenetic patterns to confer an evolutionary advantage (Luna et al.2012; Br?utigam et al.2013).
Nicotinamide (NIC), a form of vitamin B3 (niacin), is a metabolite found in all cells.It can be released from NAD in the reaction where the enzyme poly(ADP-ribose) polymerase (PARP) is involved in repairing single-strand DNA breaks, which can be caused by oxidative stress (Rissel and Peiter 2019).NAD-dependent histone deacetylases, using NAD as a substrate, also release NIC (Briggs et al.2017).Furthermore, NIC can inhibit PARP and NAD-dependent histone deacetylases, creating feedback loops (North and Verdin 2004; Rissel and Peiter 2019).Since increased histone acetylation promotes more open chromatin,NIC-inhibited deacetylation thereby has an explicit epigenetic effect, increasing histone acetylation and the resulting opportunity for transcription to occur (Avalos et al.2005).
NIC has been hypothesized to play a role in eukaryotic cells as a stress signaling molecule when it is released by PARP as a consequence of DNA strand breaks (Berglund 1994).Earlier studies showed that NIC levels increased in stressed plant tissues (Kalbin et al.1997) and that multiple defense pathways were activated by NIC (Berglund et al.1993a, 1993b, 2017).NIC has also been shown to decrease the level of DNA methylation in plant tissue cultures (Berglund et al.2017).A hypomethylating effect was also seen in spruce plants grown from treated seeds (Berglund et al.2016).
In plant cells, NIC is metabolized to nicotinic acid (NIA),which in turn is further metabolized to NAD orN-methyl nicotinic acid (trigonelline) (Noctor et al.2006).NIC is a PARP inhibitor (on protein level), but NIA and trigonelline are not.Both NIC and NIA improved stress tolerance to the heavy metals Cd, Zn and Cu inSalix viminalis(Ohlsson et al.2008) and protect against DNA damage and cell leakage caused by oxidative stress (Berglund et al.2017).
To our knowledge, there has so far been no large-scale gene expression study of NIC-treated plants.We thus explored the underlying molecular mechanisms behind NIC-induced changes in genes involved in defense and DNA methylation to form a general picture of the potential importance of NIC seed treatment for inducing stress tolerance in plants.In other studies ofinteractions between pests/pathogens and plant defense or stress response systems, leaves and needles have been the primary focus, leaving roots relatively unexplored.Here we examined changes in transcript abundances in roots from 3-month-old Norway spruce seedlings using RNA-seq technology to elucidate any changes in gene expression induced by NIC seed treatment.
Materials and methods
Plant material and seed treatment
Norway spruce seeds, collected in the orchard of ?hn (origin 57°00’ N, alt.55 m), were treated in water with 2.5 mM NIC and 1 μM Tween 80 for 4 h and gentle shaking at 23 °C in the dark.Control seeds were similarly treated in water-Tween 80.Directly after treatment, seeds were sown in wet peat and kept at 23 °C in the dark until germination started after 1 week.Plants were grown at 23 °C with an 18 h light/6 h dark cycle under fluorescent lighting tubes(150 μmol m–2s–1).After 1 month, the seedlings were transplanted in fresh peat.Seedlings were harvested 3 months after sowing, snap frozen in liquid nitrogen, then stored at-80 °C.Roots were ground into powder in liquid nitrogen before RNA extraction.
RNA extraction
RNA was extracted using the Qiagen RNeasy Plant Mini Kit and the manufacturer’s protocol.RNA integrity and quantity were measured using an Agilent 2100 Bioanalyzer.The limit for RNA integrity was RIN > 8.0.
Library construction and RNA-seq analysis
Three biological replicates were created for each of the two experimental conditions.Library preparation and RNA sequencing were carried out at National Genomics Infrastructure Sweden at SciLifeLab in Stockholm.Libraries were constructed from 200 ng total RNA each, using the Illumina TruSeq Stranded mRNA Library Preparation Kit according to the manufacturer’s instructions.Sequencing was performed with the Illumina HiSeq 2500 (Illumina,San Diego, CA, USA) using a standard protocol with read lengths of 2 × 100 bases.The average sequencing depth was 26 million reads (Table S1).The mapping frequency was 53%.The sequences are publicly available at EBI ArrayExpress (accession number E-MTAB-11007).
Data analyses
RNA-seq data were mapped using a pseudoalignment algorithm, which produces robust, accurate, and fast results compared to other strategies (Bray et al.2016), in the software Kallisto (Bray et al.2016).Kallisto uses transcripts per million (TPM), a normalized value for each transcript,making it easy to compare different samples.Differentially expressed genes (DEG) were identified using a method designed for downstream analysis of the mapped data.The algorithm takes into account the alignment statistics.It is implemented in the R package sleuth (Pimentel et al.2017).The threshold for differential expression was set top< 0.01.The reference sequence for mapping was high confidence coding sequences from theP.abiesgenome v1.0 (Nystedt et al.2013).An online platform developed by the Norway spruce genome project, congenie.org, was used to download genomic data for annotation and enrichment analysis(Sundell et al.2015).For biological interpretation of the data, the enriched DEGs were annotated for biological processes using tools available at congenie.org and the complete genome available as a reference data set.Thep-values were adjusted for multiple-testing and represent false discovery rates.Data for an epigenetic regulating subset of genes were compared to expression data for the same genes inP.abiesin other experiments and the corresponding genes inA.thalianaandPopulus trichocarpa.The data were retrieved from congenie.org, atgenie.org and popgenie.org, respectively(Sundell et al.2015).
Validation with qRT-PCR
Quantitative reverse transcription PCR (qRT-PCR) was performed for all six sequenced samples.First strand cDNA was synthesized using Superscript III according to the manufacturer’s instructions.For the qRT-PCR analyses, stress response genes CHIT4, ETR1 and MYB77, which were identified as differentially expressed in the RNA-seq analysis, were selected as suitable for the experimental conditions used in the qRT-PCR.QRT-PCR primers were designed using the online Primer3 software (Untergasser et al.2012),using default parameters except for a primer melting temperature of 60 °C and a product length of 100–150 bp.A list of the primer sequences is available in Table S2.qRT-PCR was performed in a C1000 Thermocycler (Bio-Rad) using the SYBR green kit (Bio-Rad) with a reaction volume of 10 μL.All reactions were carried out in triplicate, and a no-template control was run for each primer-pair.Actin (PaACT) was used as the reference gene (Yakovlev et al.2006).Quantification was performed using the 2-ΔΔCqmethod (Livak and Schmittgen 2001).
Results
Differentially expressed genes are associated with stress and epigenetics
Six RNA-seq libraries were generated from roots of six spruce seedlings, three from NIC-treated seeds, three control plants water-treated seeds.In the comparison of the number of sequences in the NIC-treated samples with that of the control group using a stringent cut off, 357 genes were found to be differentially expressed; 98 were upregulated and 259 downregulated (Tables S3 and S4).In the comparison of transcript abundances of up- and downregulated genes using the median tpm (transcripts per million) found in control samples (Fig.S1), the median expression of all upregulated genes was 15 TPM and 8.7 TPM for all downregulated genes.Sequence data are publicly available at EBI ArrayExpress (accession number E-MTAB-11007).
The gene ontology (GO) enrichment analysis confirmed enrichment for stress-related genes among the upregulated subset and for genes connected to epigenetic regulation among the downregulated subset (Fig.S2 A and B).A subset of differentially expressed stress- and epigeneticsrelated genes was then selected for further characterization,described next.
Upregulated stress-response genes
When all upregulated genes were screened for biological functions in stress pathways (Table 1), involvement in both abiotic and biotic stress responses was found.
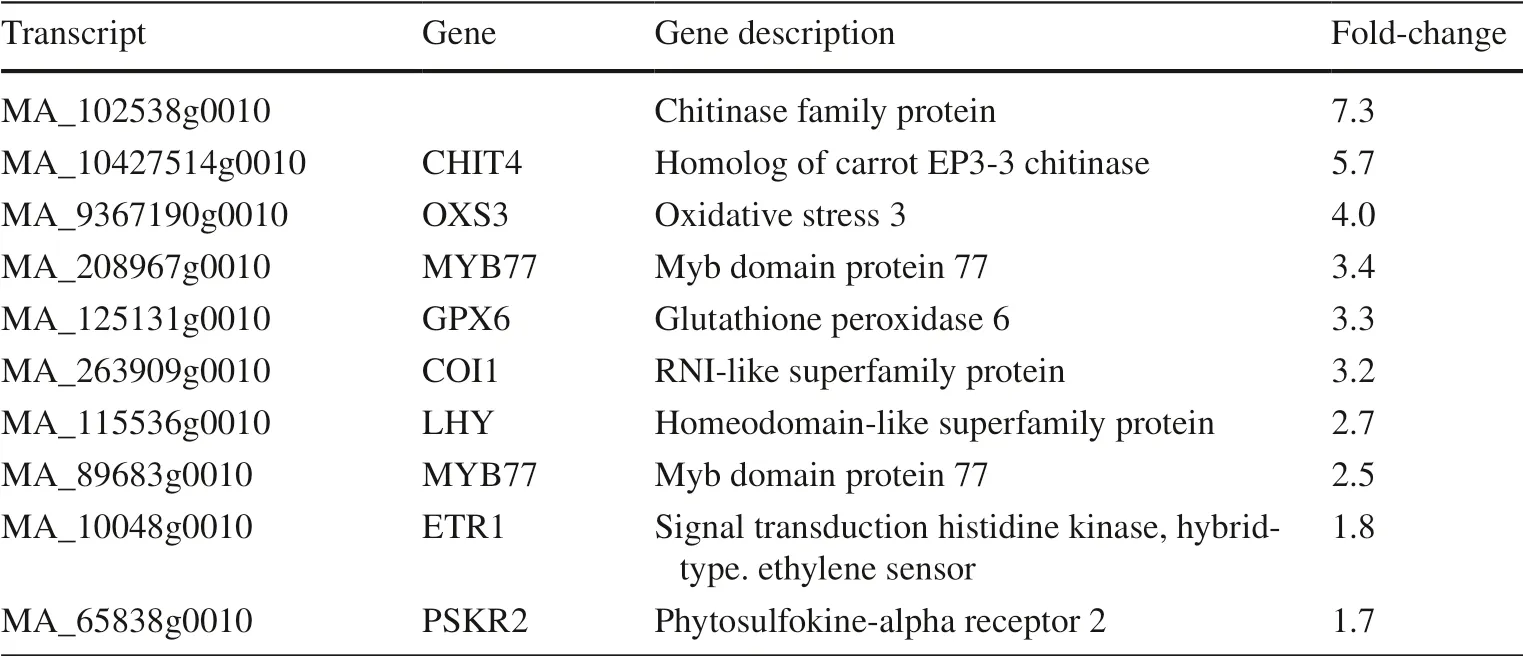
Table 1 Some of the stressrelated transcripts upregulated after NIC seed treatment
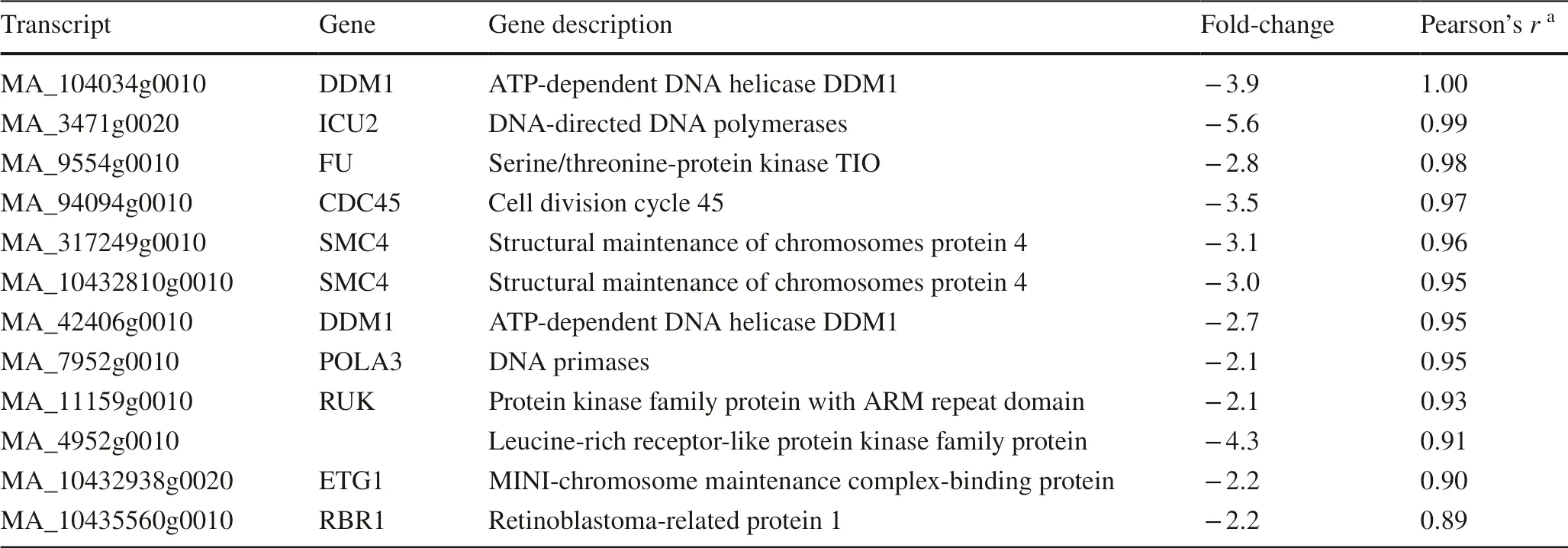
Table 2 GO annotation of epigenetic-related transcripts downregulated after NIC seed treatment
Myb transcription factors comprise one of the largest groups of transcription factors in plants.They contain a highly conserved DNA-binding domain and regulate numerous stress-related genes (Li et al.2015).In this study, two myb genes were upregulated: LHY and MYB77(MA_115536g0010, MA_89683g0010).
Gene expression of several genes involved in defense against biotic stress were induced.Chitinases play a role in pathogen defense by breaking down chitin in insect exoskeletons and fungal cell walls (Nunes and Philipps-Wiemann 2018).Two of the genes with increased transcript levels belong to the chitinase family (MA_102538g0010,MA_10427514g0010).Other upregulated pathogen defense response genes included LECRK44 (L-type lectin-domain containing receptor kinase IV.4, MA_76027g0010) and PSKR2 (phytosulfokine-alpha receptor 2, MA_65838g0010)(Loivam?ki et al.2010; Wang et al.2014a).LECRK44, a kinase in the plasma membrane, has been reported to be involved in plant immunity (Wang et al.2014b).PSKR2 is a receptor for phytosulfokine, a hormone involved in pathogen infection signaling (Zhang et al.2018).
Hormone receptors COI1 and ETR1 were also upregulated.COI1 is a mediator of wound- and jasmonate-signaling and is therefore potentially important for defense mechanisms (Devoto et al.2005).ETR1 (ethylene receptor 1) is involved in ethylene-induced abiotic and biotic stress pathways (O’Donnell et al.1996; Penninckx et al.1998).
Downregulated genes involved in epigenetic response
Twelve of the downregulated transcripts of genes are thought to be involved in epigenetic gene regulation,according to the Gene Ontology annotation (Table 2).The most recognized gene in this group is DDM1, which is an important chromosomal factor for DNA methylation(Alvarez et al.2010).
The correlation analysis of the set of downregulated genes with roles in epigenetic regulation revealed that their expression profiles were highly correlated (Fig.1,Table 2).For example, ICU2 (INCURVATA2) (Hyun et al.2013) has a Pearson correlation to DDM1 ofr> 0.99.Moreover, the correlation of these genes was observed across conditions and tissues in other experiments inP.abies(Fig.S3, congenie.org).Furthermore, this correlation was found forA.thalianaandP.trichocarpa(Fig.S4, atgenie.org, popgenie.org).
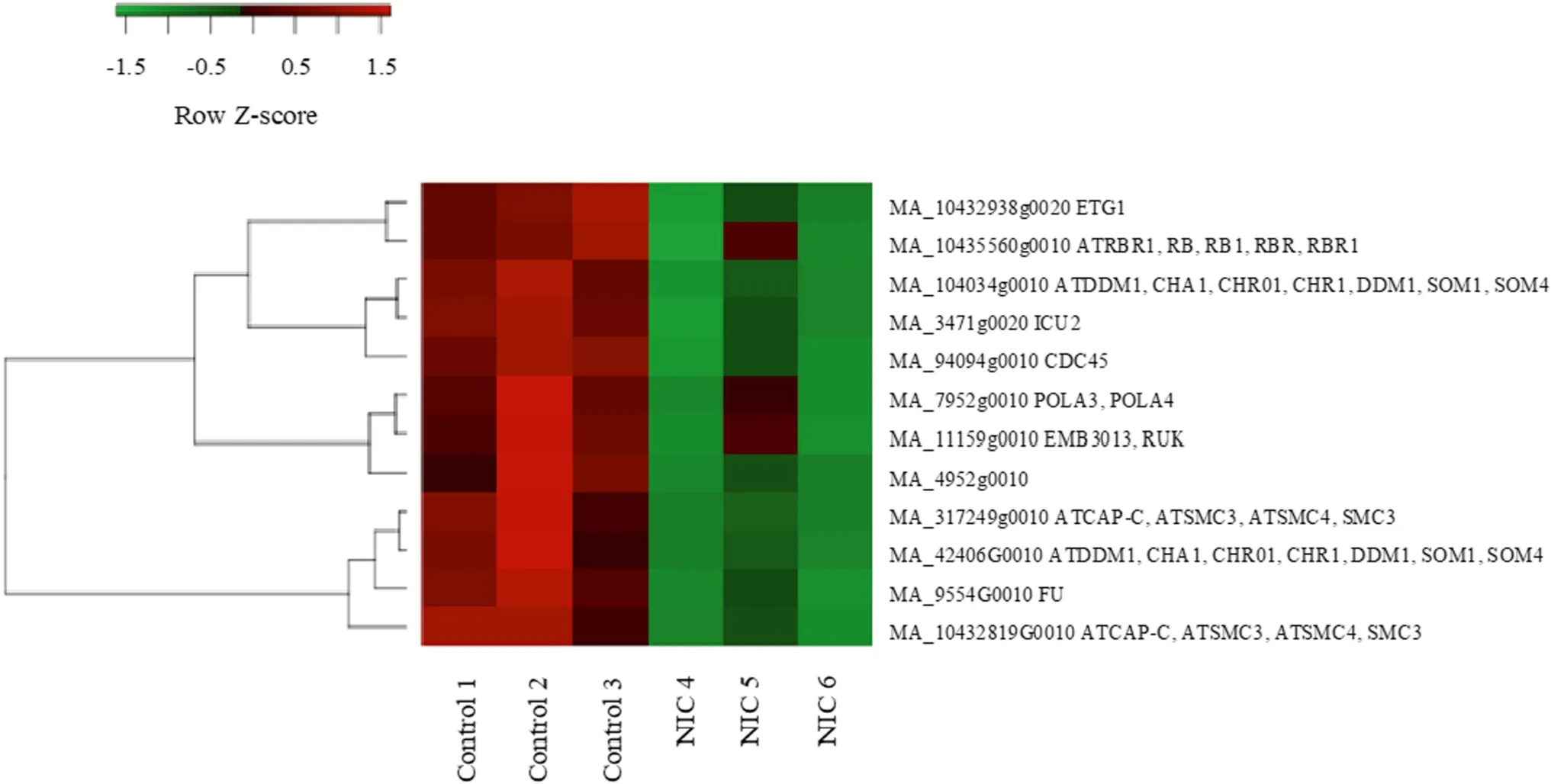
Fig.1 Coexpression of 12 NIC-responsive downregulated genes with GO annotations for epigenetic regulation
Validation with qRT-PCR supports the methodology
The qRT-PCR was performed to validate the RNA-seq expression values for three differentially expressed genes,selected because of their strong relation to stress response(see section Upregulated stress response genes) The results of the comparison between the two methods are presented in Fig.S5.The Pearson’srwas 0.91 using the gene expression measurements for each sample, respectively (Fig.S5 A).Fig.S5 B displays the differences in log2fold-changes for the tested genes after NIC treatment.This result shows that RNAseq and qRT-PCR produce concordant results.
Discussion
Long-term “memories” of stress signaling can be generated in plants through epigenetic changes to the DNA chromatin structure.In this study, we showed that treating Norway spruce seeds with NIC produces seedlings with a root gene expression profile showing several changes in expression levels of genes related to stress tolerance and epigenetics.Although analysis of epigenetic molecular hallmarks like histone modification and DNA methylation was beyond the scope of this investigation, our results indicate that NIC treatment of seeds may induce change at an epigenetic level, especially considering that our treatment at spruce embryo level (seed) is reflected in gene expression levels in months-old plants.Although further confirmation is needed, our results are in line with earlier studies that showed NIC can decrease global DNA methylation and influence an array of defense variables(Berglund et al.1993a, 1993b, 2017).The transcriptome produced by NIC seed treatment may thereby predict a stronger defense against various kinds of stress.
The plants used in this study are not clones, since we wanted to use plant material that is typically used in forestry.Therefore, the individual plants may differ genetically to some degree.Still, our analyses reveal that NIC treatment resulted in similar responses in the samples tested, which may reveal a shared mechanism underlying changes in gene expression due to stress signals generated by NIC.
Expression of genes connected to epigenetics
DNA hypomethylation in plants is associated with various stresses such as cold, heat, bacteria, insects, salinity, and wounding (Alvarez et al.2010; Santos et al.2011; Lewandowska-Gnatowska et al.2014).Logically, therefore, some genes involved in epigenetic regulation may have altered transcript abundances when stress signaling is activated.One of the downregulated genes in our study was DDM1,a key regulator of DNA methylation (Jeddeloh et al.1999),which is required for DNA methylation of transposable elements (TEs); its loss results in strong activation of TEs(Lippman et al.2004; Deniz et al.2019).TEs are frequently found near genes that are upregulated by stress in maize and other plants (Makarevitch et al.2015; Espinas et al.2016).
In addition to DNA methylation, epigenetic changes also comprise histone modification (Ramirez-Prado et al.2018).DDM1 also plays a role in histone methylation,which in turn could influence DNA methylation (Gendrel et al.2002).Another gene affecting histone modification is ICU2, which plays a key role for the maintenance of repressive histone methylation (Hyun et al.2013).Expression of ICU2 was downregulated here, as was the histone deacetylating HD2C (Histone deacetylase 2C).Downregulation of ICU2 and HD2C may lead to a chromatin state that can be reformed toward a more open configuration at some locations, allowing upregulation of certain other genes.The function of NIC as inhibitor of NAD-dependent histone deacetylases (North and Verdin 2004; Bond et al.2009) is another way for NIC to promote histone acetylation.In addition, three transcripts encoding for histones are downregulated (MA_213048g0010, MA_1527811g0010,MA_74555g0010), although the exact function of the corresponding proteins are still unknown.
Besides DNA methylation and histone modifications,the chromatin state is also stabilized by other interactions.One important protein in this regard is the histone-binding FAS1, which has been linked to transcriptional gene silencing.FAS1 is downregulated in the NIC-treated group, and its expression is highly correlated to that of DDM1 (r= 0.97).InA.thaliana, mutation of FAS1 leads to expression of TEs(Ono et al.2006).FAS1 is a subunit of chromatin assembly factor 1 (CAF-1), which cooperates with ASF1 and PCNA to contribute to chromatin assembly during replication.ASF1 and PCNA were also downregulated in this experiment and displayed a high correlation to DDM1 (r= 0.98,0.93).Downregulation of the chromatin assembly-activity of CAF-1 can lead to reprogramming of embryonic stem cells(Ishiuchi et al.2015; Kaufman 2015), and PCNA affects epigenetic inheritance (Shibahara and Stillman 1999; Zhang et al.2000).CAF-1-PCNA-mediated chromatin assembly is also linked to single-strand DNA breaks (Moggs et al.2000).PARP interacts with PCNA, which may be one mode of recruitment of PCNA to sites of DNA damage (Frouin et al.2003), but might also have other consequences for chromatin remodeling.
It can be difficult to reliably compare data obtained across multiple studies of gene expression profiles in different tissues, species, young and old plants, monocots and eudicots,all exposed to different experimental stimuli.Therefore, it is remarkable that despite the range of experimental diversity mentioned, it is clear that there is a co-variation across experiments and species between the set of downregulated genes with putative roles in epigenetic regulation that were identified in this study.High correlation and coexpression of transcripts are often indicative of a common regulating mechanism, sometimes involving motifs in the sequence of a gene or in its genomic location (Bucher 1999).In our case,NIC might have a similar kind of general epigenetic regulatory role.
Taken together, the gene expression pattern in response to NIC treatment leads to less-condensed chromatin, promoting transcription.
Expression of genes connected to stress signaling
Increased expression of certain transcription factors may confer a more-tolerant epitype in which stress adaptation is more quickly induced (López-Galiano et al.2018).In the present study, after the NIC seed treatment, the stressresponse-inducing MYB transcription factors LHY and MYB77 were upregulated in 3-month-old plants (Table 1).These transcription factors are involved in many stress-signaling pathways and can also be induced by the hormones abscisic acid (ABA), jasmonic acid (JA), salicylic acid (SA),and ethylene (ET) inA.thaliana(Shin et al.2007; Jaradat et al.2013; Grundy et al.2015).LHY regulates circadian rhythm and reactive oxygen species (ROS) homeostasis (Lai et al.2012); maximal stress tolerance with optimization of resources is achieved by circadian control.MYB77 has a role in maintaining homeostasis upon stress exposure and in the control of auxin-mediated signaling (Shin et al.2007;Jaradat et al.2013).
Many of the upregulated genes are known to specifically take part in biotic stress resistance.Examples of upregulated pathogen attack response genes are CHIT4, PSKR2, BAH1 and BAG5 (Hietala et al.2004; Doukhanina et al.2006;Yaeno and Iba 2008; Loivam?ki et al.2010; Yaqoob et al.2012) (Tables 1; S3).Chitinases can be induced both by ET and JA (Rakwal et al.2004), and two chitinase transcripts were upregulated in the present experiment.The expression of the ethylene receptor ETR1 is also upregulated after NIC treatment, which suggests that the response to ET should be faster and/or greater.ET and JA cooperatively activate stress tolerance under pathogen attack, cold, salinity, drought,and heat (Cheng et al.2013; Kazan 2015; Vos et al.2015).Furthermore, expression of COI1 (MA_263909), required for the JA response, was upregulated in our study.Another indicator that the ET/JA pathway was promoted compared to the SA pathway is that the two SA- and disease-resistance-inducing genes PAD4 and EDS1 (Rietz et al.2011)were downregulated.PAD4 and EDS1 are repressors of the ET/JA pathway inA.thaliana(Straus et al.2010).PAD4 and EDS1 are also important mediators of oxidative stress signaling (Straus et al.2010) as discussed further below.Other well-known genes involved in epigenetic processes and stress responses, such as MET1, CMT3, CMT2, ROS1 and DME (Xu et al.2018), were not differentially expressed in our study, perhaps because the seedlings analyzed were not exposed to stress as such.Rather, the seeds were treated with NIC, a putative signaling compound normally formed as a result of oxidative stress, 3 months before the analysis of the seedling roots.
Our gene expression results indicate that the ET/JA pathway is induced, whereas the SA pathway is suppressed.
Expression of genes related to oxidative stress and defense
Inhibition of PARP by exogenous chemical treatment leads to increased plant growth and to reduced production of anthocyanins and other stress-responsive molecules (Schulz et al.2012).Decreased expression of oxidative stress-related genes is also a result of PARP deficiency, as shown inA.thaliana(Vanderauwera et al.2007).In those cases, when PARP activity is inhibited, the NAD pool will be stable or increase, thus promoting growth in both stressed and unstressed plants.On the other hand, the lack of defense response observed may be due to the lack of NIC release via PARP activity, since NIC is known to stimulate several defense responses (Berglund and Ohlsson 1995).
NIC also has a general role as an antioxidant, decreasing cellular ROS content.NIC has an attenuating effect on the ROS content in human cells (Choi et al.2015; Kwak et al.2015) and increases glutathione levels in plant cells (Berglund et al.1993a, 1993b, 2017).PARP is also important for basal plant defense inA.thaliana(Briggs et al.2017).In addition, the PARP inhibitor 3-aminobenzamide prevents the increase in phenylalanine ammonia-lyase (PAL) activity caused by oxidative-stress-generating compounds (Berglund et al.1996).The transcription factor LHY, which was upregulated after NIC seed treatments in our study, is also important for oxidative stress tolerance (Lai et al.2012).Glutathione peroxidases (GPX) are involved in the antioxidant system and upregulated in response to various stresses(Rodriguez Milla et al.2003).GPX6 (MA_125131g0010)was upregulated in our experiment as was OXS3 (oxidative stress 3, MA_9367190g0010), known for its ability to increase tolerance to oxidative stress.OXS3 may operate as a chromatin remodelling factor during stress responses(Blanvillain et al.2009).ROS has an important influence on epigenetic modifications, resulting in a more open chromatin, and on stress responses (Kumar et al.2020), but the mechanisms behind these effects are largely unknown.
An important ROS-induced process in woody plants is lignification (Barros et al.2015), and genes encoding enzymes in the lignin biosynthesis pathway (e.g., cinnamyl alcohol dehydrogenase, dirigent-like protein and peroxidase)were downregulated, which may be linked to reduced ROS levels after the NIC seed treatment in our study.
Finally, there is evidence that NIC protects cells against DNA damage in yeast (R?ssl et al.2016), human (Surjana et al.2010) and plant cells (Berglund et al.2017), and we found that several DNA repair genes were downregulated in this experiment (e.g., BRCA1), which may imply that NIC reduces DNA damage.The ability of NIC to decrease oxidative stress may also partly explain the increased metal tolerance reported previously (Ohlsson et al.2008).
The changes in gene expression in response to NIC treatment suggests that NIC has an antioxidative effect.
Negative correlation between stress and epigenetic related gene expression
The expression of several of the stress-related genes discussed in the present study was strongly inversely correlated with downregulated genes that have putative roles in epigenetic regulation (Table S5).MYB77, ETR1, COI1,CHIT4, BAH1, and PSKR2 all had a Pearson’sr< –0.85 with DDM1 (MA_89683g0010).The two genes with the strongest inverse correlation (r= -0.97, -0.94) both encode membrane proteins involved in defense response.Thus, transcription of these genes might be tightly controlled by a set of potential epigenetic regulators, and/or they are affected by as yet-unidentified mechanisms, yielding an indirect relationship that is discoverable at the transcriptional level.
These data suggest that the transcription of many of the upregulated genes in this experiment is affected by a change in the abundance of putative epigenetic regulators via mechanisms that are yet to be determined.
No trade-offbetween growth and defense mechanisms induced by NIC treatment?
In experiments on the development of stress-tolerant phenotypes, there is often a trade-offbetween stress responses and growth processes.Stronger defense mechanisms come at the cost of overall growth and other function as resources are reallocated for the plant to survive the stress (Huot et al.2014).However, a positive effect of DNA hypomethylation on disease resistance inA.thalianawithout negatively affecting growth was recently demonstrated (Furci et al.2019).Similarly, we did not observe any negative effects on the growth and development of the plants originating from NIC-treated seeds.All spruce seedlings had the same size and growth rate based on visual observations and in agreement with a previous study (Berglund et al.2016).Thus,NIC may have induced a protective state that did not affect the growth rate in our experimental conditions, perhaps because there is no decrease in NAD since NIC is added exogenously and independent of PARP activity.Nevertheless, general metabolism may be slightly modified, altering plant characteristics.Thus, for each particular application,functional differences should be further explored.
The increased levels of NIC, achieved independently of PARP activity, may promote defense activation without lowering the NAD content and, consequently, without restricting growth.
Conclusions
NIC seed treatment induced changes in the transcription of approximately 350 genes in the roots of 3-month-old spruce seedlings compared to the water-treated control group.Many of the upregulated genes encoded important stress signaling molecules, which can quickly mediate a strong life-saving response in plants upon exposure to a threatening stress,while several of the downregulated genes are thought to be involved in epigenetic regulation.This study increases our knowledge about NIC-induced expression of genes involved in defense and epigenetic processes, in line with NIC as a stress-signal mediator.
AcknowledgementsThe authors acknowledge support from Science for Life Laboratory, the Knut and Alice Wallenberg Foundation, the National Genomics Infrastructure funded by the Swedish Research Council, and the Uppsala Multidisciplinary Center for Advanced Computational Science for assistance with massively parallel sequencing and access to the UPPMAX computational infrastructure.They thank Docent Henrik Aspeborg for comments on the manuscript and Docent Lauren McKee for manuscript editing.
Author contributionT.B.and A.O.conceptualized and directed the project; C.L., T.B.and A.O.designed the experiments; C.L.performed the analysis and interpreted the data, and prepared figures,tables, and references; C.L.wrote the original draft, and all authors revised the manuscript.
FundingOpen access funding provided by Royal Institute of Technology.
Open AccessThis article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source,provide a link to the Creative Commons licence, and indicate if changes were made.The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material.If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.To view a copy of this licence, visit http:// creat iveco mmons.org/ licen ses/ by/4.0/.
 Journal of Forestry Research2022年4期
Journal of Forestry Research2022年4期
- Journal of Forestry Research的其它文章
- Journal of Forestry Research
- Reversibly photochromic wood constructed by depositing microencapsulated/polydimethylsiloxane composite coating
- Surveillance of pine wilt disease by high resolution satellite
- Adaptation of pine wood nematode, Bursap helenchus xylophilus,early in its interaction with two P inus species that differ in resistance
- Pine wilt disease detection in high-resolution UAV images using object-oriented classification
- Effects of enrichmemt planting with native tree species on bacterial community structure and potential impact on Eucalyptus plantations in southern China