
Internal connections between dietary intake and gut microbiota homeostasis in disease progression of ulcerative colitis: a review
2021-06-05 06:30:10WenZengDongHeYifnXingJunyuLiuNnSuChongZhngYiWngXinhuiXing
食品科學(xué)與人類健康(英文) 2021年2期
Wen Zeng, Dong He, Yifn Xing, Junyu Liu, Nn Su, Chong Zhng,b, Yi Wng,*, Xinhui Xing,b,c,*
a MOE Key Laboratory for Industrial Biocatalysis, Institute of Biochemical Engineering, Department of Chemical Engineering, Tsinghua University, Beijing 100084, China
b Centre for Synthetic and Systems Biology, Tsinghua University, Beijing 100084, China
c Institute of Biopharmaceutical and Health Engineering, Tsinghua Shenzhen International Graduate School, Shenzhen 518055, China
ABSTRACT
Ulcerative colitis (UC) is a chronic systematic inflammation disorder with increasing incidence, unknown pathogenesis, limited drug treatment, and abundant medical expenses. Dietary intake, as a daily indispensable environment factor, is closely related to UC pathogenesis and prevention. The underlying interactions between dietary intake and UC progression are implicated with the modulation of gut microbiome as well as microbial metabolites, suggesting the complex and systematic characteristics of UC. However, the triangular relationships with dietary intake, gut microbiota homeostasis, and UC have not been well summarized so far.Here we review the recent studies of dietary intake on the regulation of gut microbiome homeostasis as well as modulation of UC progression. These findings suggest that varieties in dietary patterns result in the production of diverse microbial fermentation metabolites, which contribute to gut microbiome homeostasis through multiple manipulations including immune modulation, inflammation restriction as well as epithelial barrier maintenance, thus finally determine the fate of UC progression and give implications for functional food development for prevention and treatment of UC patients.
Keywords:
Ulcerative colitis
Dietary intake
Gut microbiome
Metabolite
Homeostasis
1. Introduction
Ulcerative colitis (UC) is a chronic, idiopathic inflammatory bowel disease (IBD) with relapsing and remitting course of superficial mucosal inflammation limited to part or all of the colon [1-3]. In clinic, UC patients usually suffer from bloody diarrhea, abdominal pain, low-grade fever, as well as malnutrition [4,5]. The quality of life in UC patients is fairly poor with disability, complications, and even an elevated risk of colorectal cancer [6,7]. With urbanization and modernization of people’s daily life, the incidence and prevalence of UC are increasing rapidly in worldwide [8]. The highest incidence and prevalence of UC have been reported in developed countries including Europe, Canada, the United States, and Australia [9]. Developing countries, especially newly industrialized countries in Asia, South America, and Africa, are facing a promptly increase in UC incidence although these countries have a lower total number of UC patients compared with developed countries [10-12]. In the medical treatment of UC, the initial goal focuses on the induction and maintenance of the disease remission, while the ultimate goal focuses on ameliorating disability, preventing colectomy, and minimizing the risk of colorectal cancer [13]. Anti-inflammatory drugs, such as 5-aminosalicaytes and steroids, are widely adopted in mild to moderately active UC, while immune suppressors and biological therapies are adopted in moderate to severely active UC [14,15]. However, the drug therapy in UC treatment is challenged by poor tolerance and distinct side-effects,which results in nearly 30% of UC patients undergo surgical treatment in their lifetime [16]. Meanwhile, extensive medical care leads to a significant economic burden in UC treatment. Previous studies reported that the annual cost was $15 020 in UC patients versus$4 982 in other patients, while lifetime total cost was $405 496 in UC patients versus $209 697 in other patients [17,18]. The annual medical expenditure for IBD treatment in Europe and the United States was estimated at €12.5-29.1 billion and $8.1-14.9 billion, respectively,suggesting the importance and urgency to discover new approaches in the investigation and treatment as well as prevention of UC [19].
Until now, there is no clear conclusion in the pathogenesis of UC, which is caused by multifactorial influences for UC including genetic, immune, environment, and microbiome. [20]. The relapsing chronic inflammation in UC is partially the results of an imbalance in immune response [21]. Dendritic cells, macrophages, and helper T cells promote pro-inflammatory response, while regulatory T cells restrain over-activated immune response and are closely associated with UC remission [22,23]. Meanwhile, genetic variation is also implicated with immune pathogenesis of UC. Up to now, 200 genomic loci are reported to relate with IBD [24,25], in which loci implicate with regulatory pathway, cell migration, and epithelial barrier function are more specific to UC [26,27]. Environmental factors including urbanization, antibiotics intake, and digestive tract infection elevate the susceptibility of patients to UC, while smoking is negatively related with the susceptibility to UC [28,29]. Moreover,negative mental states affected by major life stressors, anxiety, and depression are also related with increased susceptibility to UC.Physiological stress can promote intestinal inflammation through nervous system and, leads to UC exacerbation [30,31]. Dietary intake,as a daily indispensable environmental factor, plays a critical role in UC development and prevention. From the moment of birth,first diet intake of breast milk is correlated with ameliorated susceptibility to childhood UC among the newborns [32]. Accumulating in vivo studies observed that diverse dietary patterns are closely related to UC progression. Dietary intake of fresh fruits, fibers, olive oil, and amino acids are implicated with UC remission and decreased risk,while high-fat diet, high-sugar diet, and processed foods intake exacerbated colonic inflammation as well as UC progression [33-36].Clinical trials have found that the specific carbohydrate diet, antiinflammatory diet, and semi-vegetarian diet ameliorated the disease activity and lead to the disease remission in UC patients [37,38].
Of note, dietary intake largely shapes gut microbiome of the hosts, which is considered to be an important endocrine organ and plays a pivotal role in UC pathogenesis [39]. Recent findings have advanced our insight into the interplay between gut microbiome and UC progression through the regulation of immune response,inflammation, and epithelial barrier [40]. A lack of early childhood exposure to microbes is associated with impaired maturation of host immune response, causing elevated susceptibility to UC [41].The dysbiosis of gut microbiome stimulates content of potential pathogens including Escherichia coli, leads to increased expression of lipopolysaccharide and further aggravates chronic inflammation as well as UC progression [42]. Furthermore, several mucolytic microbes including Ruminococcus gnavus and Akkermansia muciniphila are related with the impaired mucosal barrier functions as well as chronic colitis symptoms. With the advancement of UC investigation, the relationships between diet and UC as well as between microbiome and UC have been reviewed. In brief, on the one hand, omic studies revealed the internal connections between gut microbiome and other pathological factors of UC and figured out that the gut microbiome was shaped by host genetic and environmental factors including diet,public health, antibiotics, and others. In turn, it interacted with the enterocytes, impacted on the immune systems, and regulated mucosal barrier integrity, thus further modulated UC progression [43,44]. On the other hand, recent reviews indicated that the dietary intake of macronutrients involving carbohydrate, protein, fat, and others largely contributed to UC susceptibility. Meanwhile, dietary therapies for IBD treatment have been widely studied as it reshapes our dietary patterns and may alter the exposure to the deleterious substances such as some specific food additives and refined sugars, thereby affecting the host immune systems and the disease progression [45,46], therefore, gut microbiome as well as the dietary intakes play important roles in UC progression. However, little is summarized about the triangular relationships with dietary intake, gut microbiome homeostasis and UC progression as well as the insights into functional food development for UC prevention [47-49].
In this review, we would highlight the internal connections between dietary intake and gut microbiome homeostasis in UC progression, which gives insight into the comprehensive understanding of dietary intake as well as microbiome in UC pathogenesis. Our work also provides the comprehensive perspective for innovation and mechanism investigation of dietary therapy and prevention of UC patients.
2. Intrinsic links between UC and gut microbiome
2.1 Microbiome change in the pathogenesis of UC
Colonic microbiome is dominantly comprised of bacteria, most of which are Firmicutes and Bacteroidetes, while less abundant of Proteobacteria, Actinobacteria, and Verrucomicrobia coexist in healthy individuals [50]. The homeostasis of gut microbiome is closely related with human wellness, whereas the disturbance and disruption of colonic microbiome are relevant to disease progression of systematic disorders including asthma, type I diabetes, as well as UC [51]. Clinical trials revealed that the dysbiosis of gut microbiome was accompanied with decreased microbiome diversity, ameliorated content of Firmicutes, as well as elevated content of Proteobacteria in UC patients. In addition, depleted key species including Verrucomicrobia, Roseburia, Akkermansia were found, while the excess of pathogenic species containing Streptococcus,Bifidobacterium and Enterococcus were reported in UC patients[52,53]. The structural shift of gut microbiome decreased content of the resident gastrointestinal bacteria and ameliorated stability of micro-ecological environment in colonic lumen.Therefore, it gave opportunities to potential pathogens in the invasion of epithelial barrier, and further perpetuated chronic inflammation in UC progression.
Despite the individual difference of gut microbiome, numerous human studies have exerted a unified trend of gut microbiome change in UC patients, which was summarized in Table 1. The elevated population of potential pathogens, which mainly belong to Proteobacteria and are Gram-negative, including adherent-invasive E. coli, Escherichia-Shigella, and Helicobacter, played a critical role in UC pathogenesis [23]. The presence of adherent-invasive E. coliled to the invasion of colonic epithelium as well as the stimulation of immune response, whileEscherichia-Shigellaiwas responsible for UC relapse [54]. As a potential pathogenic entity,Helicobactercould result in persistent inflammation in colonic epithelia. Other studies also found that increased levels ofRhodococcusandBacteriodes fragiliswere correlated with aggravated inflammation flare and disease progression in UC patients.
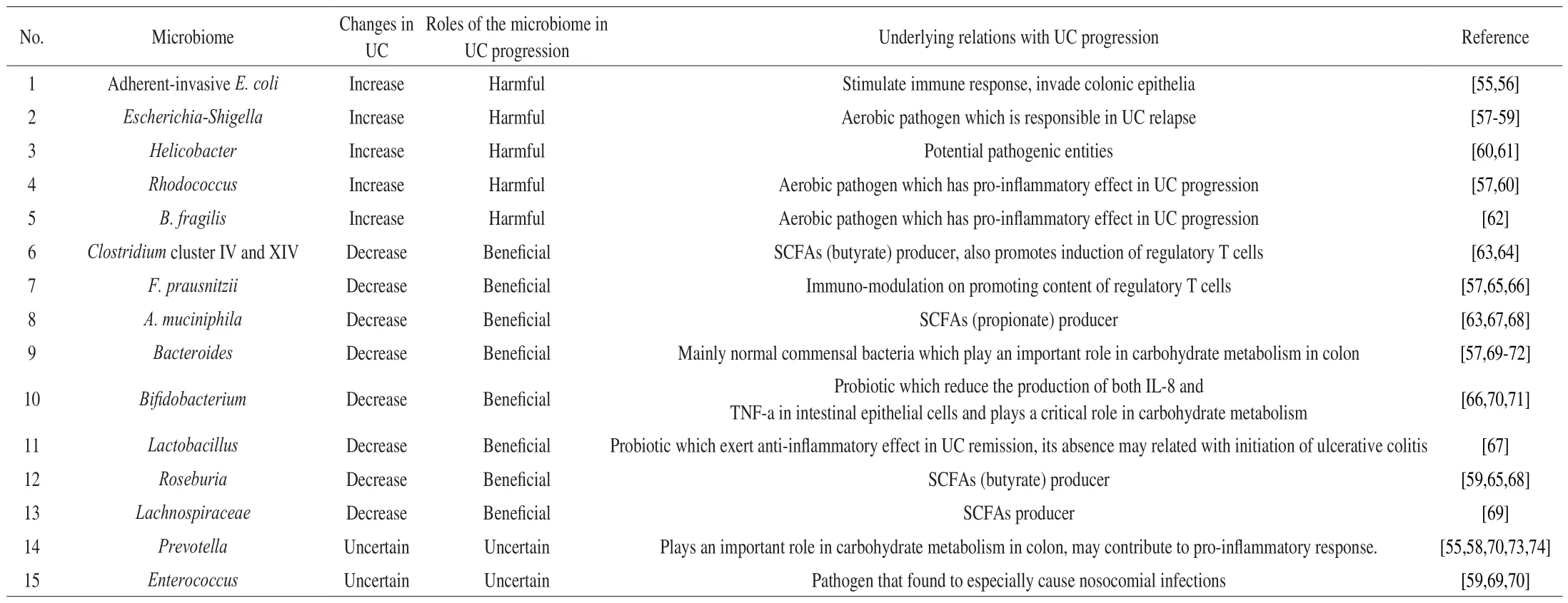
Table 1Gut microbiome change and its relations in UC patients.
In comparison with harmful pathogens, reduced levels of certain bacteria, especiallyFaecalibacterium prausnitziias well asClostridiumcluster IV and XIV, were closely implicated in UC remission. As one of the richest commensal bacteria in healthy individuals,F. prausnitziiexerted protective effect through stimulating regulatory T cells and promoting secretion of antiinflammatory cytokines in UC remission [75].Clostridiumcluster IV and XIV largely contributed to microbiome diversity and possessed abilities to produce butyrate which was a momentous category of short chain fatty acids (SCFAs) that resulted from regulatory T cells and exerted anti-inflammatory effect [76]. Meanwhile, decreased levels of other SCFAs producers includingRoseburia, Lachnospiraceae,andA. muciniphilawere also found in UC patients, indicating that the imbalance between pro-inflammatory and anti-inflammatory response contributed to persistent chronic inflammation [68]. Some studies also revealed the regulatory effect of probiotics in UC remission.The absence ofLactobacillusmight be related with initiation of UC,while the presence ofBifidobacteriumreduced the production of both interleukin (IL)-8 and tumor necrosis factor-α (TNF-α) in intestinal epithelial cells and played a critical role in carbohydrate metabolism,suggesting their protective role of beneficial microbiome through regulating T cells and promoting the secretion of anti-inflammatory cytokines in UC pathogenesis [77]. Other microbiomes includingPrevotellaandEnterococcuswere also implicated with intestinal infection as well as pro-inflammatory response in UC progression,whereas the content of which fluctuates case by case, suggesting the complexity of human gut microbiome.
2.2 Microbial metabolites are associated with UC progression
Since the presence of gut microbiome interferes gut homeostasis and disease progression, microbial metabolites link the gut microbiome and hosts together in UC progression and participate in the regulation of immune and metabolic responses (Table 2). For instance, butyrate in SCFAs, which was produced from microbial fermentation of indigestible carbohydrates and proteins, could be utilized as a nutrient supplement for enterocytes [78]. SCFAs also inhibited pro-inflammatory cytokine secretion in macrophages and stimulated suppression activity of regulatory T cell (Treg) through G-coupled protein receptor (GPR) including GPR41, GPR43, and GPR109 [79]. Whereas SCFAs production was negatively correlated with disease progression in UC patients, suggesting the beneficial roles of SCFAs through multiple manipulations in gut homeostasis.Indole compounds, derived from the metabolism of green vegetables and tryptophan byLactobacillusandAllobaculum, were commonly suggested as the ligands for endogenous aryl hydrocarbon receptor(AhR). The stimulation of AhR signaling promoted IL-22 secretion as well as the production of antimicrobial peptide and tight junction proteins, indicating the protective role of indole compounds in mucosal barrier healing [80,81]. The metabolism of other indigestible components containing sulfur as well as nitrogen induced sulfide and amines production, respectively. The presence of amines in colon mainly came from the fermentation of proteins and promotes infiltration of inflammation in gastrointestinal tract, thus correlated with UC progression [82]. Hydrogen sulfide (H2S), considered as the third gasotransmitter and involved in the regulation of inflammation and motility, was produced by sulfate-reducing bacteria in gut lumen and is positively correlated in UC patients [83]. H2S degraded mucus layer through the reduction of disulfide bridge in mucins and induced energy deficit by the inhibition of butyrate oxidation, thus impaired epithelial barrier integrity and aggravated micro-ecological environment in colon [84]. Besides the metabolites mentioned above, recent studies revealed other metabolites including phenols,salicylic acid, and others were also implicated in UC progression [85].However, how to correlate metabolites with specific source microbiomes and identify their roles in UC progression still remains difficult and needs further investigation.

Table 2Microbiome metabolites in relations of UC.
3. Dietary intake modulates gut microbiota homeostasis and influence disease progression of UC
3.1 Probiotics
Probiotics are living organisms that can bring benefits when administrated into the hosts. Dietary intake of probiotics could manipulate the gut microbiota as well as exerting the immunemodulatory, antimicrobial and anti-inflammatory activities in digestive tract [92,93]. Accumulating evidences have suggested that the probiotics play a pivotal roles in the regulation of gut flora and alleviation of UC progression [94,95]. The most applied probiotics wereLactobacilliandBifidobacterium, whileBacillus,Streptpcoccus,and probiotic-rich foods were also documented.
Inin vivostudies of UC, dextran sulfate sodium (DSS) and 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced murine colitis model were the most applied chemical-induced colitis models with simplicity and high success rate [96]. DSS-induced murine colitis could resemble human UC in the aspects of histology and immunology with the defects of high reagent expense and individual difference [97]. Although TNBS-induced murine colitis could overcome the defects, TNBS performed colitis was largely dependent on the mouse strains and was more likely to exert Crohn’s disease(CD)-like symptoms [98].
In DSS and TNBS induced colitis model mice, the administration of probiotics could restore the gut microbiome homeostasis through the increasing microbiome diversity, ameliorating potential pathogens,and improving structural composition of resident microbiomes in colitis mice [99,100]. Dietary intake ofBifidobacteriuminduced UC remission and microbiome change within 3 days [101]. Recent studies adopting KEGG pathway annotation found that dietary intake ofBacillus amyloliquefacienschanged the pathways related with carbohydrate, lipid and amino acid metabolism in colitis mice [102]. Despite chemical-induced colitis, aging and obesity were closely correlated with the chronic inflammation with an elevated susceptibility of colitis pattern in colon [103,104]. Recent studies using high-fat diet induced colitis mice revealed that the administration of probiotics restored the gut microbiome homeostasis in the same way as the chemical-induced colitis model. Of note,immune modulation on Th17 differentiation and inhibition of inflammation related with NF-κB and mitogen-activated protein kinase (MAPK) signaling pathways were found in obese mice with dietary probiotics. Moreover, ameliorated obesity and liver damage were also observed, suggesting the systematic amendment in the treatment of high-fat induced colitis [105-107]. In the age-dependent colitis model, early studies found that adopting probiotic mixture IRT5 andLactobacillus pentosuselevated levels of Verrcomicrobiota and Prevotellaceae, respectively. Ameliorated MAPK, mechanistic target of rapamycin (mTOR) as well as forkhead box O3 (FOXO3)signaling pathway were observed in probiotic treatment mice,which indicated the inhibition of over-activated inflammatory response [108,109]. The positive regulatory effect of dietary probiotics on gut homeostasis and UC remission was also verified in IBD dogs and UC patients [110].
These studies suggested that in the aspects of gut microbiome,dietary intake of probiotics could quickly lead to the structural change, improve microbiome diversity and restore homeostasis through ameliorating potential pathogens includingE. coli, restoring the content of resident gastrointestinal bacterial speciesBacteroidesand recovering Firmicutes/Bacteroidetes (F/B) ratio as well as Proteobacteria/Bacteroidetes(P/B) ratio. In the aspects of disease progression, dietary intake of probiotics led to UC remission in a systematic manner through inhibiting inflammation-related pathways,modulating immune cell differentiation as well as protecting colonic epithelial barrier integrity.
3.2 Dietary proteins and amino acids
Recent reviews highlighted the interactions between protein and gut flora. Dietary intake of protein led to complex proteolytic processes in digestive tract, which further resulted in the production of SCFAs, branch-chained fatty acids, ammonia, indoles and so on [91].These complex interactions could modulate the disease progression of UC in positive or negative directions.
Recent studies found that the amount of protein intake had a significant impact on the gut microbiome as well as UC progression.Medium protein diet (30%) led to disease remission while high protein diet (53%) exacerbated the UC progression. High protein diet led to an increase inPorphyromonas,Ethanoligenens, andHaemophiluswhile a decrease inBacteroide, which indicated the deterioration of microbiome composition. Medium protein diet led to an increase in Eubacteriaceae (Firmicutes) and Bifidobacteriaceae (Actinobacteria)as well as butyrate-producing genera throughout the resolution phase, which indicated the positive impact of gut microbiota on UC remission [111]. In addition to the amount of protein intake, the types of protein intake also showed different effects on UC progression.Recent studies focusing on animal protein diet and plant protein diet revealed that the excessive intake of animal protein aggravated DSS-induced colitis in mice with an increase of monocytes and iNOS+cells. The over-activated immune response as well as inflammation were related with the ameliorated diversity of bacteria and fungus and led to increased relative abundance of genera Enterococcus,Streptococcus, Turicibater and Escherichia. Meanwhile, excessive intake of plant protein did not influence UC progression, families Bifidobacteriaceae and Desulfovibrionaceae were enriched in plantbased protein diet. The difference in digestibility of dietary protein might result in discrepant biological functions and interactions between gut microbiome [112]. Other studies adopting red meat intake and high protein diet also observed similar results on gut microbiome and disease progression in both normal mice and DSS-induced colitis mice [113,114]. Early study using cheese whey protein observed disease remission with anti-inflammation activity and increased fecal mucin secretion in DSS-induced colitis rats. Increased content of Lactobacilli and Bifidobacteria were found by cheese whey protein intake. As cheese whey protein is rich in threonine and cysteine, which is the limiting factor in mucin synthesis, dietary intake of threonine and cysteine also exerted similar effect than cheese whey protein [115]. Some studies adopting dietary amino acids also found the similar benefits in UC treatment. Dietary intake of arginine restored microbial diversity as well as increased abundance of Bacteroidetes and decreased abundance of Verrucomicrobia[116]. Dietary intake of tryptophan changed serum amino acid profile, alleviated increasing levels of Asp, Met, and Pro induced by acetic acid. Although there is no change in microbiome diversity,dietary intake of tryptophan ameliorated the relative abundance of Clostridiaceae at the family level and Candidatus, Clostridium,Coprococcus at the genus level compared with colitis group [117].
These studies suggested that proper intake of plant-based protein,dairy protein and amino acid were beneficial to UC remission. The potential connections were protective microbial metabolites including SCFAs and indole compounds that restored microbiome environment and regulate immune response. Meanwhile, excessive intake of dietary protein, especially animal-based protein, might be correlated with H2S and amine production, thus further exacerbated UC progression with over-activation of immune response and increased contents of potential pathogens.
3.3 Dietary fats
High-fat diet was positively correlated with prevalence and incidence of UC. Numerous evidence suggested that high-fat diet could promote chronic inflammation, colitis symptom, as well as UC exacerbation in vivo [118-120]. On the one hand, high-fat diet aggravated UC progression through disruption of gut epithelial barrier and promotion of inflammation, which resulted of elevated secretion in pro-inflammatory cytokines, stimulation of immune response, absence of tight junction proteins, and decline of secretory cells including paneth cells and goblet cells [121,122]. On the other hand, high-fat diet disrupted the gut microbiome homeostasis with ameliorated microbiome diversity and elevated P/B ratio and F/B ratio [123]. Moreover, inflammation-associated microbiota including Peptostreptococcaceae and Streptococcus were increased and SCFAs producer Prevotellaceae was decreased in high-fat diet mice [124],elevated abundance of pathobionts including Atopobium and Proteobacteria were also reported [121]. In addition, maternal high-fat diet also perturbed gut microbiome homeostasis in murine offspring, led to impaired gut barrier function and elevated susceptibility to UC, this influence retained in murine offspring for at least 5 weeks, suggesting the long-term effect of high-fat diet on gut microbiome [124].
Despite the quantity of dietary fat intake, early studies observed that the type of dietary fat was also important in the development of murine colitis. Monounsaturated fatty acid (MUFA) rich diet including olive oil ameliorated inflammation in Citrobacter rodentium induced colitis model through the induction of intestinal alkaline phosphatase which detoxifies lipopolysaccharide. Omega-6 polyunsaturated fatty acids (n-6 PUFA) rich diet including corn oil and saturated fatty acids(SFA) rich diet including milk fat did not exert better remission [125].Further investigation on gut microbiome revealed that, although all of the high-fat diets similarly resulted in increased content of Firmicutes,there were differences in family level of the gut composition which could shape the core function of gut community and further regulate UC progression. An n-6 PUFA rich diet enhanced levels of Turicibacteraceae, which resulted in a more invasive microbiome with increased cell motility and virulence. MUFA rich diet increased abundance of Clostridiaceae and Peptostreptococcaceae, which resulted in microbiome with increased metabolic activities. SFA rich diet promoted contents of Erysipelotrichales, which resulted in a microbiome with increased capacity for energy harvest [126].
These findings suggested that the quantity and the type of dietary fat regulated gut flora composition and further affected UC progression. High-fat diet aggravated microbiome dysbiosis and led to colonic inflammation and exacerbated UC. The negative effect of fat intake might be implicated with cell metabolism, metabolites including prostaglandin E2and 5-lipoxygenase exerted proinflammatory effect in UC progression [127]. Meanwhile, the type of dietary fat also shaped the gutflora and regulated UC progression.
3.4 Dietary carbohydrates
Dietary intake of carbohydrates provides energy for human body and composes basic structure components in living cells. Also, it is implicated in UC progression and influences gut microbiome in various ways [128,129]. According to the utility rate in human body,it could be divided into digestible carbohydrates and non-digestible carbohydrates.
High-sugar diet, which mainly contains digestible carbohydrates,was closely related with an elevated risk of UC [8]. Recent studies on high-sugar diet revealed its negative impact on UC progression in vivo. In 10 days investigation, 50% sucrose of high-sugar diet aggravated colonic inflammation and epithelial barrier permeability in DSS-induced colitis mice. High-sugar diet induced an elevated level of Verrucomicrobia and decreased levels of Firmicutes, which suggested an increased susceptibility to colitis and gut microbiome dysbiosis. Ameliorated levels of Lachnospiraceae family were accompanied by depletion of SCFAs in colon [130]. These findings suggested the negative impact of dietary sugar in microbiome regulation and UC progression.
Besides digestible carbohydrates like sugars, numerous studies were focusing on non-digestible carbohydrates in UC treatment.Early study using konjac oligosaccharide significantly ameliorated TNBS-induced murine colitis, oligosaccharide treatment alleviated abundance of E. coli as well as Enterococcus and elevated the content of probiotics including Lactobacillus and Bifidobacterium,thus resulted in restored SCFAs production in gut lumen [131]. A 12-weeks in vivo study using HLA-B27 transgenic rats validated the protective effect of non-digestible oligosaccharides. Dietary intake of fructo-oligosaccharides increased contents of Bifidobacteria and Enterobacteriaceae, whereas the content of Clostridium cluster IV is decreased [132]. Early study adopting resistant starch in DSS-induced colitis mice ameliorated disease activity and promoted colonic epithelial integrity, the downtrend in potential pathogens including A. muciniphila, Enterococcus spp., and E. coli were observed [114].Early study focusing on dietary fiber and inflammation process in DSS-induced colitis mice revealed the relationship between gut microbiota and inflammation response. Compared with zero-fiber diet group, Porphyromonadaceae, Rikenellaceae, and Lachnospiraceae were found predominantly in high fiber diet group. These changes in gut microbiota led to production of SCFAs and activation of NOD-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome pathways through G-coupled proteins GPR43 and GPR109a, resulted in disease remission in DSS induced colitis [133]. Recent study by Marten’s group [134] examined the protective effect of dietary fiber in gnotobiotic mice with characterized human gut microbiome. Mice feeding with fiber-free diet resulted infiber depletion in digestive tract and stimulated content of mucus-degrading microbiome including Bacteroides thetaiotaomicron and Bacteroides caccae. Administration of fiber-rich diet maintained the balance between fiber-degrading microbiome and mucus-degrading microbiome, thus sustained gut homeostasis and mucus layer integrity. Fiber-degrading microbiomes like Bacteroides ovatus and Eubacterium rectale were abundant in fiber-rich diet mice. Further investigation adopting mucosal pathogen C. rodentium led to lethal colitis in fiber-free mice, dietary intake of fiber ameliorated weight loss and colonic epithelial inflammation in fiber-rich mice.
Although the above studies confirmed the positive regulation of dietary fibers on the gut microbiome and UC disease remission in mice, low carbohydrate intake was preferred in human trails. Among them, specific carbohydrate diet (SCD) has received more attention about IBD treatment in recent years. Initially adopted in the treatment of celiac disease, this special diet does not contain any complex carbohydrates including grains and some vegetables [135,136]. In a 12-weeks human trial, SCD diet reduced the content of Escherichia,Haemophilus, Clostridium, Streptococcus, Proteobacteria, and Bacteroides, inverted the ratio of Bacteroides to Firmicutes in active IBD patients. Along with the remission in gut microbiota, SCD diet led to disease remission in both UC and CD patients [137]. Similarly,recent human study on fermentable carbohydrates (FODMAPs)revealed the negative impact of FODMAPs including fructans,galacto-oligosaccharides, and sorbitol, on active UC patients. Dietary intake of FODMAPs exacerbated the severity of pain, bloating,flatulence and faecal urgency in UC patients, compared with placebo(glucose) [138]. Further study on quiescent UC patients adopting low FODMAP diet disclosed disease relief and increased quality of life scores compared with control group. Although there were no changes in inflammation and gut microbiome diversity, lesser contents of Bifidobacterium adolescentis, Bifidobacterium longum, and F. prausnitzii were found in low FODMAP diet group [139].
In a word, in vivo studies revealed that restricted sugar intake and promoting dietary intake of non-digestible fiber could sustain gut microbiome homeostasis through regulation of mucin-degradation microbiomes and SCFAs producing microbiome, thus regulated the immune response and inflammation process in UC. However,human studies revealed low carbohydrate diet is preferred in both active and quiescent UC patients with adequate relief and no obvious improvement among inflammation. This contradiction phenomenon could be the result of gut microenvironment differences between human and mice. Meanwhile, complications including irritable bowel syndrome and celiac disease in UC could also regulate gut microbiome as well as disease progression in UC patients, suggesting the complexity in UC pathogenesis. The underlying mechanism of dietary carbohydrate in UC treatment needs further investigation.
3.5 Other dietary patterns
Modern dietary patterns including high salt intake and processed foods intake were also correlated with exacerbation on chronic inflammation and UC progression [140]. Recent studies revealed the negative impact of high-salt diet in murine colitis model. High salt intake reduced content of probiotic Lactobacillus while increased levels of Lachnospiraceae and Oscillospira in gut microbiome,the disruption of microbiome resulted in aggravated DSS-induced colitis, elevated p38/MAPK signaling pathway and inhibited butyrate production [141,142]. Dietary intake of emulsifiers, detergentlike molecules that adopted in processed foods, changed gut microbiome composition with decreased levels of Bacteroidales and increased level of mucolytic operational taxonomic units(OTUs) including R. gnavus and A. muciniphila. Disrupted mucus layer and chronic inflammation in colon epithelia were found in germ-free mice received fecal microbiota from dietary emulsifier mice, which indicated elevated susceptibility to UC [40,143].Dietary intake of titanium dioxide nanoparticles, another additive widely adopted in food industry, also aggravated DSS-induced colitis and immune response in vivo. The negative effect of nanoparticles was highly correlated with nanoparticle size and changes in gut microbiome. It might be related with the disturbed gut microbiome diversity and reduced abundance of probiotics including Bifidobacterium and Lactobacillus [144]. In addition, recent studies revealed that dietary change also elevated susceptibility to UC and exacerbated colitis symptoms in vivo. Despite the overall composition of different murine chow diet showed no significant difference, the diet switch of daily foods resulted in the expansion of Proteobacteria and Verrucomicrobia, while decreased abundant of Bacteroidetes was found. This change aggravated UC progression, although the reason why diet switch led to microbiota change remains unknown [145].Another in vivo study applying over 40 unique dietary combinations suggested that macronutrients including protein and fiber had the greatest impact on DSS-induced murine colitis, while dietary patterns containing casein, isolated soy protein, and psyllium largely modulated UC progression through microbiota-dependent manner [146].Another study adopting fasting-mimicking diet found lowcalorie and low-protein diet stimulated Allobaculum and probiotic Bifidobacteriaceae abundance in DSS-induced colitis mice, and led to UC remission with ameliorated inflammation as well as increased colonic stem cells [147]. Early study adopting high-fat and sucrose western diet found western diet could affect gene expression as well as survival of Lactobacillus plantarum, and ameliorated L. plantarum activity with disrupted gut flora homeostasis, thus aggravated UC progression [148].
Besides modern dietary patterns, the dietary intake of functional components involving polyphenols, indole derivatives, and other prebiotics also largely shaped gut microbiome, thus further modulated UC progression. Intriguingly, accumulating evidences suggested that the dietary intake of fresh fruits and vegetables containing large amount of vitamins, polyphenols, and polysaccharides was beneficial to alleviating chronic inflammation as well as UC progression[30,149]. Recent studies adopting polyphenol-rich mango beverage in DSS-induced colitis rats found that the dietary intake of mango beverage led to an increase in the fecal content of L. plantarum,Lactococcus lactis, and SCFAs-producer Clostridium butyrium. The amelioration of colitis might be achieved through HDAC1/AMPK/LC3 axis which affected by butyrate and polyphenol metabolites [150].Another studies using polyphenol-rich bronze tomato also found structural change of fecal microbiome with significant decrease in F/B ratio and significant increase in Parabacteroides, A. muciniphila, and Lactobacilli, compared with normal tomatoes that did not contain polyphenols. Moreover, anti-inflammation effect of bronze tomato on IL17A and interferon γ (IFN γ) production further alleviated UC progression in DSS-induced colitis mice [151,152]. Recent studies focusing on dietary berries, which were rich in diverse vitamins and polyphenols, including strawberry, cranberry, and aronia berry found the anti-inflammatory and immuno-modulatory effect in colitis mice.Intriguingly, these studies showed similar impact on maintaining gut microbiome homeostasis and rescuing colitis symptom through promoting microbiome diversity, increasing levels of probiotics containing Lactobacillus and Bifidobacterium, and ameliorating conditional pathogenic bacterium covering Akkermansia, Dorea, and Bilophila. These changes in gut flora were correlated with restored production of SCFAs, alleviated secretion of pro-inflammatory cytokines, and inhibited activity of NF-κB and MAPK signaling pathways, thus further led to UC remission [153-155]. Recent studies found Pleurotus eryngii, as the homology of medicine and foods, also exerted protective effect in gutflora manipulation and UC remission.Increased levels of Adlercreutzia and Akkermansia while decreased levels of Desulfovibrionaceae were found in dietary P. eryngii group[156]. Another study found dietary intake of Hericium erinaceus could sustain gut microbiome homeostasis through the regulation between potentially pro-inflammatory microbiomes and potentially anti-inflammatory microbiomes. Polysaccharide components of H. erinaceus might play a prebiotic role while the alcoholic soluble components of H. erinaceus exerted bactericidin-like and immunomodulatory effects in murine colitis model, suggesting the multiple modulation effect of dietary H. erinaceus in UC treatment[157]. Dietary intake of propyl-propane thiosulfonate form garlic restored microbiome diversity and alleviated F/B ratio in DSS-induced colitis mice, ameliorated MAPK signaling activity as well as inflammation pattern were found [158]. These studies suggested that dietary intake of different fruits and vegetables could modify gut microbiome variously. Of note, the functional components found in fresh fruits and vegetables might have shaped the dietary patterns and the gut microbiome, while the microbial metabolites and derivatives of functional components might largely contribute to UC progression.Meanwhile, multiple manipulations of gutflora coexisted in dietary fruits and vegetables which contain diverse types of biological components, thus led to UC remission.
4. Conclusion and prospect
In this review, we summarized the interplay between food intake and gut microbiome homeostasis in UC progression. Dietary intake could rapidly fluctuate dynamic homeostasis of gut microbiome within a day, while long-term administration of specific diet modulates UC progression, at least in part, through interactions with microbiome and production of microbial fermentation metabolites.Dietary intake of probiotics directly influences the gut microbiome composition and improves the microbial environment, which is beneficial to UC remission. The quantity as well as quality of protein intake determine the production of diverse microbial metabolites including indole compounds, hydrogen sulfide and amines, thus further modulate UC progression in positive or negative ways. Different dietary fats could induce various microbiome changes upon the saturation index of fats, while cell metabolism of dietary fats might contribute to colonic inflammation in UC progression. In vivo studies have suggested that the fermentation of indigestible carbohydrates provides abundant SCFAs and nutrients for enterocytes as well as gut microbiome, hence plays a positive role in UC remission. Dietary fruits and vegetables control gut microbiome and UC progression through a more compositive way as these foods contain multiple bioactive components.
These findings have suggested the diverse manipulation and complex mechanism of dietary intake in the modulation of gut microbiome homeostasis in UC progression (Fig. 1). Dietary therapy is becoming a potential method in UC treatment compared with drug therapy and surgical treatment [159]. Intestinal microbiome as well as microbial metabolites are connecting links between dietary intake and UC progression. With the development of microbialbased therapies including fecal microbial transplantation, the mutagenesis and screening of gut microbiome, which are beneficial to UC remission, could be a potential choice in UC treatment.Meanwhile, dietary intake of fermented foods were implicated in UC remission. The fermentation process of numerous indigestible dietary ingredients result in the production of diverse functional bioactive components that could lead to UC remission. Latest findings have observed the protective role of dietary functional components derived from probiotics fermentation of fruits, vegetables, and grains in UC prevention [160,161]. Long-term administration of functional fermentation components could modulate micro-ecological environment as well as microbial composition in colonic lumen,thus further restrains UC progression through a safe, convenient, and enduring manner. These findings have indicated a potential direction about the development as well as investigation of functional foods in UC prevention and treatment in future. Still, there are increasing differences between in vivo studies and clinical trials of dietary intake in UC treatment. UC is a class of systematic gastrointestinal disorder and the gut microbiome has compact and complex crosstalk with the host in the aspects of immune system, gut micro-environment,and genetic variation. Although diverse dietary patterns largely modulate the gut microbiome which plays an indispensable role in UC progression, it is still hard to define the core microbiome that changes specifically in UC due to the insufficient data and confusing results [162]. The correlation between specific gut microbiome and UC progression is still challenging and largely remains unknown,therefore, the comprehensive understanding of gut microbiome as well as dietary intake in UC pathogenesis is further needed.
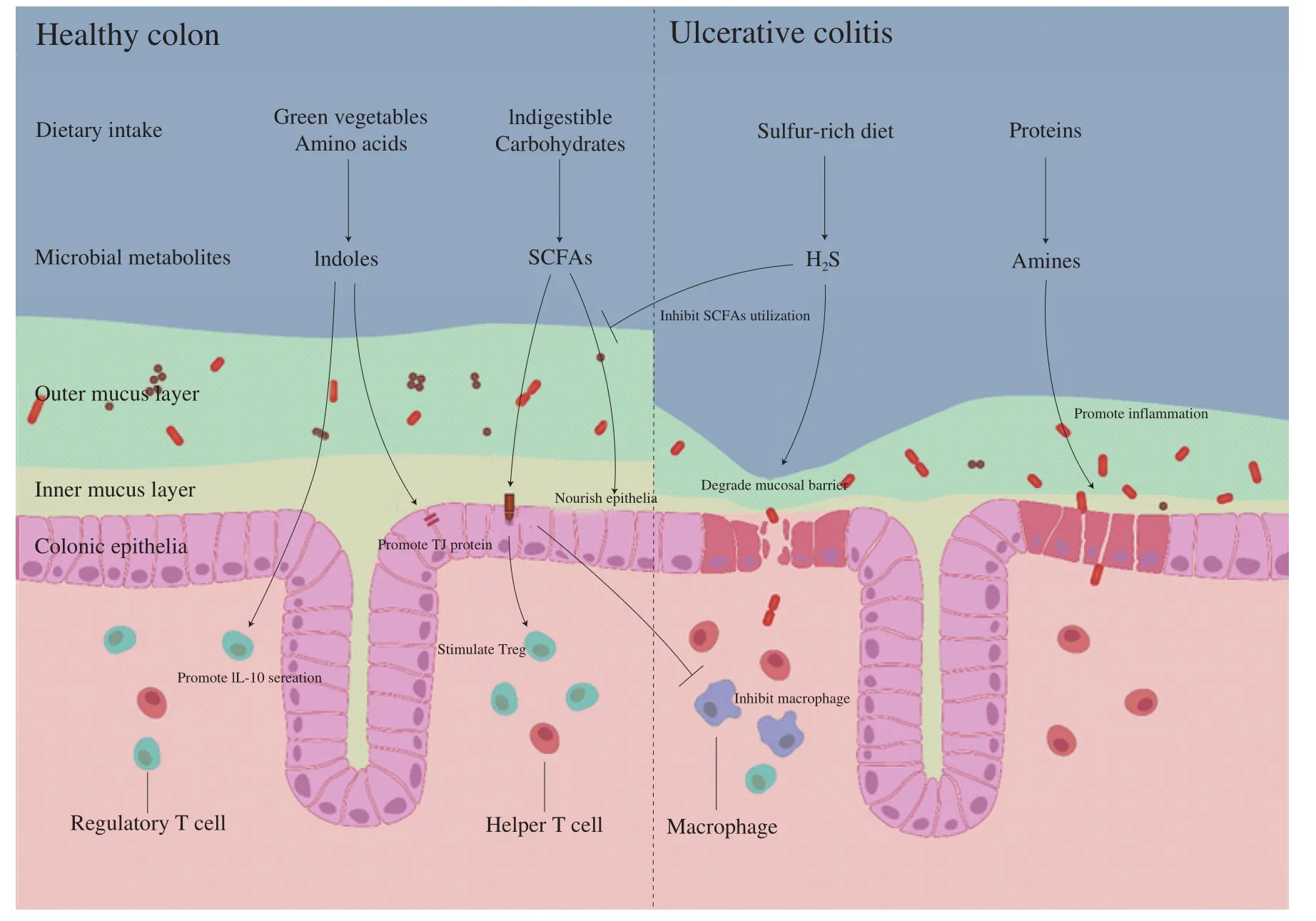
Fig. 1 The underlying connections of dietary intake and microbiome homeostasis in UC progression.
Conflict of Interest
The authors declared that they have no conflicts of interest to this work.
Acknowledgements
This work was supported by the National Key Research and Development Plan, China (2016YFD0400203-4).

- 食品科學(xué)與人類健康(英文)的其它文章
- Predominant yeasts in Chinese Dong fermented pork (Nanx Wudl) and their aroma-producing properties in fermented sausage condition
- Isolation and identification of fungi found in contaminated fermented milk and antifungal activity of vanillin
- The association between folic acid supplementation, maternal folate during pregnancy and intelligence development in infants: a prospective cohort study
- Investigation of physicochemical, microbiological and sensorial properties for organic and conventional retail chicken meat
- Preparation and physicochemical/antimicrobial characteristics of asparagus cellulose films containing quercetin
- Simultaneous determination of 74 pesticide residues in Panax notoginseng by QuEChERS coupled with gas chromatography tandem mass spectrometry