
Thermal Biology of Cold-climate Distributed Heilongjiang Grass Lizard,Takydromus amurensis
2020-12-30 06:59:26XinHAOShiangTAOYuMENGJingyangLIULuoxinCUIWanliLIUBaojunSUNPengLIUandWengeZHAO
Asian Herpetological Research 2020年4期
Xin HAO,Shiang TAO,Yu MENG,Jingyang LIU,Luoxin CUI,Wanli LIU,Baojun SUN,Peng LIU*,# and Wenge ZHAO*,#
1 College of Life Science and Technology,Harbin Normal University,Harbin 150025,Heilongjiang,China
2 Key Laboratory of Animal Ecology and Conservation Biology,Institute of Zoology,Chinese Academy of Sciences,Beijing 100101,China
3 College of Chemistry and Life sciences,Zhejiang Normal University,Jinhua 321004, Zhejiang,China
4 School of Chemical Engineering,University of Science and Technology Liaoning,Anshan 114051,Liaoning,China
Abstract Thermal biology traits reflect thermal adaptations to an environment and can be used to infer responses to climate warming in animal species.Within a widespread genus or species,assessing the latitudinal or altitudinal gradient of thermal physiological traits is essential to reveal thermal adaptations and determine future vulnerability to climate warming geographically.We determined the thermal biology traits of a cold-climate distributed liza rd,Takydromus amurensis,a nd integra ted published thermal biology traits within the genus Takydromus to reveal a preliminary geographical pattern in thermal adaptation.The mean selected body temperature (cloaca temperature; Tsel),critical thermal maximum (CTmax),critical thermal minimum(CTmin),and optimal temperature for locomotion (i.e.,sprint speed; Topt) of T.amurensis were 32.6,45.1,3.1,and 33.4 °C,respectively.The resting metabolic rates of T.amurensis were positively related to temperature from 18 °C to 38 °C.We compared the traits of tropical T.sexlineatus,subtropical T.septentrionalis,and T.wolteri with T.amurensis and found that the CTmax and thermal tolerance range (the difference between CTmax and CTmin; TTR) increased toward high latitudes,whereas CTmin increased toward low latitudes in these four Takydromus lizards.According to this preliminary pattern,we speculate the species at medium and low latitudes would be more vulnerable to extreme heat events caused by ongoing climate warming.We highlight the importance of integrating thermal biology traits along geographical clues,and its potential contribution to evaluate the vulnerabilities of species in the context of climate warming.
Keywords counter gradient,CTmax,CTmin,thermal biological trait,thermal tolerance range,Tsel,Takydromus
1.Introduction
Ongoing climate warming has imposed novel stressors on animals (e.g.,Thomaset al.,2004),which has affected their populations in a number of ways.For example,many species have altered their distributions,and some populations face risk of collapse (e.g.,Poundset al.,1999;Wilsonet al.,2005).Recent studies have shown that the thermal safety margin (difference between environmental temperature and critical maximum temperature,CTmax) of low-latitude animals is narrower than that of animals living in mid and high latitudes,and the thermal biological characteristics of low-latitude animals are more susceptible to temperature variation,indicating that low-latitude animals are more vulnerable to climate warming (e.g.,Dillonet al.,2010; Güsewellet al.,2017;Sundayet al.,2014).Thus,a detailed understanding of the thermal biology traits and the thermal responses of animals is not only useful for studying adaptations to thermal environments,but also necessary for determining the vulnerability to thermal variation caused by climate warming.
Thermal biology traits have been shown to be important proxies of the thermal adaptation of animals to their environments,as measured by selected body temperatures (Tsel),CTmax,CTmin,and the thermal dependency of performance such as locomotion (e.g.,Huey and Hertz,1984; Huey and Kingsolver,1989; Angillettaet al.,2002a).Unlike endotherms,which can maintain a stable body temperature in a wide temperature range at great energetic costs (Bennett and Ruben,1979),the body temperatures of reptiles and other ectotherms are regulated by thermal interactions with the environment and thus are more susceptible to thermal variations and their effects upon behavior and physiological processes(Huey and Stevenson,1979; Angillettaet al.,2002a,2002b).Accordingly,reptiles have a high risk of extinction owing to climate warming (e.g.,Sinervoet al.,2010).In addition,they are affected by internal and external factors (Wanget al.,2017).Through the analysis of the thermal biology traits of widespread genera or species,inferences can be made regarding the geographical patterns and interspecific variation of traits.This can aid in determining the vulnerabilities of widespread genera to climate change along geographical gradients (Hueyet al.,2009; Sinervoet al.,2010).
Reptiles can maintain their body temperature within a moderate range by thermoregulation,with a range of optimal body temperatures for different processes(Van Dammeet al.,1991; Bauwenset al.,1995).The selected body temperatures of a reptile represent the body temperatures without biotic or abiotic constraits(Hueyet al.,1979; Hertzet al.,1993),which can be determined by the temperature gradient in the laboratory,although extrapolation of laboratory data to field populations should be interpreted cautiously.Because Tselrepresents the “ideal” range of body temperatures ectothermic animals aim to maintain through behavioral thermoregulation,it is normally different from active body temperatures (Lichtet al.,1966; Avery,1978).CTmaxand CTminare the maximum and minimum temperatures at which vitality can be sustained,respectively (Hertzet al.,1983; Gilbert and Miles,2017).If the body temperature is lower than CTminor higher than CTmax,reptiles would lose motor function and suffer cold narcosis (temperature lower than CTmin) or muscle spasms (temperature higher than CTmax) (Lutterschmidt and Hutchison,1997;Angilletta,2009; Camacho and Rusch,2017).Therefore,CTmaxand CTminare used to determine the viability of reptiles in various thermal environments (Hertzet al.,1983; Deutschet al.,2008).
Metabolism is the primary physiological process that determines an organism’s demand from the environment and the allocation of energy among body functions(Brownet al.,2004).In ectotherms,the resting metabolic rate is often used as a proxy for metabolism and is defined as the energy expenditure of animals while in the resting,fasting,and nonproductive states (Andrews and Pough,1985; Christianet al.,1999; Berget al.,2017).In reptiles,resting metabolic rates are highly sensitive to body temperature,requiring the organism to respond quickly to variations in the thermal environment (Maet al.,2018a,2018b; Sunet al.,2018).As another critical function,locomotion is also significantly sensitive to body temperature and is related to fitness because of its importance for foraging,avoiding predators,and choosing mating partners (Bauwenset al.,1995; Chenet al.,2003; Sunet al.,2018).In summary,evaluating the thermal sensitivity of critical indices (i.e.,metabolic rates and locomotion) is essential to understand thermal adaptation in ectotherms (Angilletta,2009).
Takydromusliza rds have a la rge geographical distribution,with a significant latitudinal span,ranging from cold-temperate to tropical regions.As a species that inhabits cold climates,the Heilongjiang grass lizard,Takydromus amurensis,is a small lacertid lizard (up to 70 mm snout-vent length [SVL]) which is distributed in northeast China and the boundary of China,Russia,and the Korean Peninsula (Xuet al.,2017).Like otherTakydromuslizards in mainland China,T.amurensisinhabits areas at the boundary of forests and grasslands or scrub habitats.Females are able to lay one or two clutches from May to July,with an average clutch size of six eggs (Zhaoet al.,1999).T.amurensisis distributed in high latitudes,which typically exhibit low average temperatures and drastic thermal fluctuations (Figure 1).Thermal biology traits should be investigated inT.amurensisto obtain a comprehensive understanding of thermal adaptation in cold-climate ectotherms.The thermal biology of several other congener species has been previously studied,includingT.wolteri,T.septentrionalis,andT.sexlineatus,which inhabit subtropical and tropical areas,respectively (Jiet al.,1996a; Chenet al.,2003; Zhang and Ji,2004).By assessing the same traits inT.amurensis,i.e.,those investigated in these previous studies,we can improve our understanding of the specific variations and preliminary patterns of thermal biology traits inTakydromusspecies along latitudinal and altitudinal gradients.
Figure 1 Annual temperature environments for Anshan population of T.amurensis,where we collected the lizards.The red line indicates the average air temperature along the year,and grey area indicates the daily fluctuations of air temperature.
We determined selected body temperatures,thermal tolerance (i.e.,CTmaxand CTmin),and thermally sensitive traits including resting metabolic rate and sprint speed as focal study traits for the Heilongjiang grass lizard,T.amurensis.We aimed to (1) contribute with new data on a cold-climate distributedTakydromusspecies (i.e.,T.amurensis) for the first time; (2) compare thermal biology traits amongTakydromuslizards with published data concerningT.septentrionalis,T.wolteri,andT.sexlineatus;and (3)summarize preliminary patterns of variation in Tsel,CTmax,CTmin,and optimal body temperature for performance inTakydromuslizards along a latitudinal or altitudinal gradient.
2.Materials and Methods
2.1.Study species collectionWe collected adultT.amurensis(n=13; 4 males and 9 females) in late April 2016,using either a noose or by hand,from Anshan (see details in Figure 1 for annual thermal environments),Liaoning,China (41°01’ N,120°7’ E;~210 m).After collection,we transported the lizards back to the laboratory in Beijing.The lizards were individually housed in plastic terraria(350 mm × 250 mm × 220 mm,length × width × height).The terraria were set up with a mixture of moist soil and sand,with randomly placed patches of grass.The lizards were housed in a temperature-controlled room at 18°C under a natural photoperiod,with a supplementary heating lamp suspended above one end of the terrarium from 06:00 to 20:00.During the hea ting period,temperatures in the terraria ranged from 18 °C to 40 °C.Food (crickets and larvalTenebrio molitor) and water were providedad libitum.After three days of captivity,thermal biology traits were started to be determined.
2.2.Selected body temperatures (Tsel)Selected body temperatures (Tsel) were determined according to the established protocol,with minor modifications (Shuet al.,2010).In brief,Tselwas measured in a custom-made terrarium (1000 mm × 500 mm × 300 mm,length × width× height) placed in a temperature-controlled room at 18 °C.The terrarium was fitted with grass substrate and‘natural’ retreats to mimic the natural environment.One heating light bulb (275 W) was suspended above one end of the terrarium,creating a thermal gradient from 18 °C to 60 °C during the heating period.The heating period lasted 14 h daily from 06:00 to 20:00.On the first day,a group of lizards (four to five) were introduced into the terrarium from the cool end at 16:00 for acclimation to the surroundings.Then,on the second day,the body temperatures of the lizards were measured twice at 09:00 and 15:00,with an UNT-325 electronic thermometers(UNT T-325,Shanghai,China) by inserting the tip of the probe into cloacas.During the measurements,each lizard was captured by hand and its body temperature was measured immediately (within 30 s),without disturbing the other lizards in the terrarium.Then,the average of the two body temperature records of each lizard was estimated and used as the lizard’s selected body temperature.
2.3.Thermal toleranceThe critical thermal minimum(CTmin) a nd critical thermal maximum (CTmax)temperatures of the lizards were determined in a programmed incubator (KB 240,Binder,Germany).We cooled (for CTmindetermination) or heated (for CTmaxdetermination) lizards from a starting temperature of 28 °C at a rate of 1 °C per 10 min.During the cooling and heating,we continuously monitored the behavior of the lizards.When the lizards lost the capacity to respond to intense stimulation and could not right themselves after being turned over,the body temperatures were recorded as CTminor CTmax,respectively (Zhang and Ji,2004; Xu and Ji,2006).The lizards were then moved to a 28 °C chamber to recover.The protocol stands that if a lizard cannot recover from the cold or heat shock,its record is eliminated for further analysis (Liet al.,2017).However,in this study all the lizards were recovered from the shock
2.4.Locomotor performanceLocomotor performance was estimated using sprint speed.The sprint speed was measured at five test temperatures ranging from 18 °C to 38 °C (18 °C,23 °C,28 °C,33 °C,and 38 °C),based on the reported range of field body temperatures of the species(Xuet al.,2017),in a randomized sequence.Locomotor performance was measured once per day under one temperature treatment (18 °C,33 °C,28 °C,23 °C and 38 °C),in a sequence order.Before the test,the lizards were placed in an incubator at the test temperature for approximately 2 h for acclimation.To ensure that body temperatures of lizards matched those of the respective test treatment,we measured the body temperatures of a subset of lizards before starting stimulating them to run(Sunet al.,2014).Locomotor performance was tested by stimulating the lizard to run through a racetrack,which was recorded by an HD video camera (Sony,DCRSR220E,Japan).The racetrack was 1500 mm × 100 mm× 150 mm,with intervals marked every 250 mm.Each lizard was stimulated by a paintbrush to run twice at each temperature with an interval of 1 h for rest.The videos were analyzed by Windows Movie Maker.For each lizard,the fastest speed through 250 mm for each time was recorded,and the average of the two fastest records was used as the sprint speed (Sunet al.,2014).
2.5.Resting metabolic rate (RMR)The resting metabolic rates were estimated by respiratory gas exchange rates measured at five random test temperatures (18 °C,23 °C,28°C,33 °C,and 38 °C).Before the test,the lizards were fasted for at least 12 h.All lizards were acclimated at the correspondent test temperature for 2 h in an incubator (KB 240,Binder,Germany).Then,the lizard was enclosed within the respirometry chamber placed in the incubator (KB 240,Binder,Germany).The respiratory gas exchange was determined using a closedflow respirometry system with a volume of 281.4 mL(Stable System International Inc.Las Vegas,NV,USA),and the resting metabolic rate was estimated via the CO2production rate using a previously established method(Sunet al.,2018).For stabilizing the gas composition of the system (i.e.,baseline),we opened the system to the air for approximately 5 min before changing the system to closed-circuit respirometry.Then,the carbon dioxide production rates (i.e.,VCO2) in the closed circuit were continuously recorded for approximately 10 min.To minimize the effects of circadian rhythms,measurements were conducted from 10:00 to 18:00.The metabolic rates were calculated as the CO2production per gram of body mass per hour (mL/g/ h),following the equation metabolic rate=VCO2× volume/body mass,whereVCO2is the CO2production rate in percentage (%/h) in the closed circuit with a volume of 281.4 mL.
2.6.Statistical analysisThe normality of distributions and homogeneity of variance were tested using the Kolmogorov-Smirnov test and Bartlett’s test prior to analysis.First,we tested for sex differences in Tsel,CTmax,and CTminusing one-way ANOVA,with sex included as a factor.As there were no significant differences (allP> 0.05),we pooled the data of both sexes for all these traits.For the analysis of locomotion,we performed the Gaussian (R2=0.473),modified Gaussian (R2=0.523),Lorentzian (R2=0.513),Weibull (R2=0.515),and Pseudo-Voigt regressions (R2=0.512) to analyze the sprint speed to body temperature,respectively.With the best fitting,the modified Gaussian regression was employed to analyze the sprint speed,with the following equation:Sprint speed=y0+ae ^ {-0.5 [(Tb-b)/c]2}; whereTbis body temperature; a,b,c andy0are parameters; and e is the natural constant.This was used to calculate the optimal temperature for sprint speed.Repeated measures ANOVAs were used to analyze the effect of test temperature and sex on resting metabolic rate and sprint speed.
Data of previously illustrated traits ofT.septentrionalisfrom Zhoushan,Zhejiang (29°32’-31°04’N,121°30’-123°25’E;~200 m),T.wolterifrom Chuzhou,Anhui(32°15’-32°21’ N,118°07’-118°18’ E;~260 m),andT.sexlineatusfrom Shaoguan,Guangdong (24°50’ N,113°30’ E;~290 m) (Jiet al.,1996a; Chenet al.,2003;Zhang and Ji,2004; unpublished data) were collected from published figures with Plot Digitizer (http://plotdigitizer.sourceforge.net/).The thermal traits ofT.septentrionalis(collected throughout May and July),T.wolteri(collected in May),andT.sexlineatus(collected in mid-April) were obtained from the literature.Only data collected during the reproductive season were selected to ensure comparability with the results of the current study(Jiet al.,1996a; Chenet al.,2003; Zhang and Ji,2004).General linear models were fitted to assess the latitudinal tendency ofTsel,CTmax,CTmin,and Toptfor sprint speed ofTakydromuslizards.As the four species inhabit similar altitudes (i.e.,from~200 m to~290 m),we did not include this variable in the model.The fact that elevation is fixed in the design of the study provides clarity to be able to detect the effect of latitude.
3.Results
The Tsel,CTmin,and CTmaxforT.amurensiswere 32.55 ±0.46 °C,3.06 ± 0.13 ℃,and 45.10 ± 0.11℃,respectively(see details in Table 1).The sprint speed was significantly affected by body temperature,increasing with body temperature from 18 °C to around 33 °C and then decreasing at higher temperatures (F4,48=5.495,P<0.0001; Figure 2a).The optimal body temperature forT.amurensisthat maximized sprint speed was 33.20 °C according to the modified Gaussian regression.Resting metabolic rate was significantly positively related to temperature from 18 °C to 38 °C (F4,48=40.708,P<0.0001; Figure 2b).The Sex had no effect on the resting metabolic rate or sprint speed (bothP> 0.05).
In the fourTakydromusspecies,Tselwere similar,as indicated by the non-significant tendency along latitudes(y=32.99-0.55x,R2=0.37,P=0.38; Figure 3a).When we integrated the published thermal tolerance data from otherTakydromuslizards,we observed that the CTmaxvalues (y=45.43-0.93x,R2=0.78,P=0.11) remained similar; however,CTmindecreased as latitude increased(y=2.137+1.04x,R2=0.95,P=0.02) (Figure 3b),indicating that the thermal tolerance range increased with latitude.
Figure 2 Locomotor performance (sprint speed) (a) and resting metabolic rate (RMR) (b) of T.amurensis at different temperatures.RMR was expressed as CO2 production per gram body mass per hour (mL/g/h).Data are expressed as mean ± SEM.
Table 1 Snout-vent length,body mass,selected body temperatures,the critical thermal maximum,critical thermal minimum and thermal tolerance range (TTR) of Takydromus amurensis.The sample size for all traits are equally 13.
4.Discussion
In the context of climate change,understanding the thermal biology traits of animals is fundamental to determining organismal vulnerabilities (e.g.,Sundayet al.,2011,2012,2014).In the current study,the thermal biology traits ofT.amurensiswere quantified at the beginning of the reproductive season,to facilitate a comparison with available data of congeners collected using similar methods.The Tsel,CTmin,CTmax,and sprint speed among different species of the genusTakydromuswere shown to be compa ra ble a nd ecologically meaningful (Jiet al.,1995,1996a; Duet al.,2000; Zhang and Ji,2004).Since the four populations live at similar latitudes (i.e.,ranged from~200 m to~290 m),we only tested the effects of latitude.
Figure 3 Tsel (a) and CTmax and CTmin (b) of four Takydromus lizards.Data are expressed as mean ± SEM.Species are listed from high to low latitude in X axis.Data are collected by Plot Digitizer(http://plotdigitizer.sourceforge.net/),from the figures and available data of references (Ji et al.,1996a; Xu and Ji,2006; Zhang and Ji,2004).
Tselis the body temperature at which biological processes function at an optimal/suboptimal level (Van Dammeet al.,1991; Hertzet al.,1993; Blouin-Demerset al.,2000; Angillettaet al.,2002a; Ortega and Martín-Vallejo,2019).InT.amurensis,the optimal temperature for sprint speed was similar to Tsel(33.20 °Cvs.32.55°C),indicating thatT.amurensiscan accurately regulate its body temperature to perform optimal/suboptimal functions.However,as we did not measure operative tempera ture (Te) a nd active body tempera tures(Ta),we were unable to determine the efficiency of thermoregulation.Numerous reptiles have been reported to have temporal,spatial,and individual variation in Tsel(Gatten Jr,1974; Andrews,1998; Stellatelliet al.,2018;Refsnideret al.,2019).There is also increasing evidence demonstrating that thermal biology traits vary among seasons (e.g.,Yanget al.,2008; Sunet al.,2014; see details in below).However,Tsel is normally only determined during the reproductive season and assumed representing the Tsel of the given species or population (Jiet al.,1995,1996a; Shine and Madsen,1996; ?orovi? and Crnobrnja-Isailovi?,2018; Refsnideret al.,2019).It should be noted that althoughT.amurensisis a cold-climate species,the TselinT.amurensisis very similar to published data forT.wolteri,T.septentrionalis,andT.sexlineatus(Figure 3a).Additionally,the lizardsZootoca vivipara,Lacerta agilis,andIberolacerta bonnaliwhich also inhabit cold climates,use thermoregulation to maintain their body temperature warm in responding to cold climates (Herczeget al.,2003;Yanget al.,2015;Ortega et al.,2016).Influence of the thermal environment and activity patterns could induce variations in Tsel(Jiet al.,1996a,1996b; Duet al.,2000;Zhang and Ji,2004).For example,lizards in forested habitats are likely to have lower Tselthan those using more open habitats.The TselofSphenomorphus indicus,whose habitats are forested,was as low as 25.7℃ (Jiet al.,1996b).On the contrary,the lizards using more open habitats and employing basking behaviors would exhibit higher selected body temperatures (e.g.,Liet al.,2017;Wanget al.,2019).T.amurensisis usually active in open microhabitat,with shuttling between open and shaded patches for thermoregulation (Zhao,2002),which may plausibly explain high TselofT.amurensis.Admittedly,we conducted selected body temperatures measurements ofT.amurensiswith established method according to those used in otherTakydromusspecies (e.g.,Jiet al.,1995;Zhang and Ji,2004),increasing literatures indicate that the selected body temperatures of ectotherms should be determined with multiple repeated measurenets (i.e.,eight to ten measurements per individual) (e.g.,Hertzet al.,1993; Liet al.,2017).We also encourage further researches should employ multiple measurements to make a better understanding of thermal preferences in lizards,includingTakydromulizards.
The thermal tolerance range increased toward high latitudes inTakydromuslizards,with relative conservative CTmaxand decreased CTmintowards high latitudes.(Figure 3b).Microhabitat usage can significantly influence CTmaxand CTmin.For example,CTmaxwas lower inS.indicus,which uses shaded habitats likely to be cooler (37.6 ℃;Jiet al.,1996b) than species occupying open habitats,such asEumeces elegans(41.9 ℃; Duet al.,2000) andT.septentrionalis(42.3 ℃; Jiet al.,1996a).Among sympatric lizards,Eremias argus(45.8 ℃) andE.multiocellta(45.1 °C)that occupying filtered microhabitats,had lower CTmaxthanPhrynocephalus przewalskii(47.1 °C),which uses open habitats (Liet al.,2017; Wanget al.,2019).Analogously,CTminwas noticeably greater in lizards that use open habitats.The CTminofE.chinensis(6.3 °C; Jiet al.,1995) andE.elegans(9.3 ℃; Duet al.,2000),species occupying open habitats,were greater than species using more spatially complex habitats,such asE.argus(1.0 ℃; Luoet al.,2006).Although all the four species compared in the current study use open habitats (Zhaoet al.,1999),T.amurensismay spend more time on open habitats in response to lower temperature conditions,as happens in other highlatitude distributed lizards (Sundayet al.,2011; Sundayet al.,2012).This activity pattern may result in increased time invested in thermoregulation (i.e.,basking) and thus result in higher CTmax.Based on this assumption,global meta-analyses have demonstrated that CTmaxincreases with latitude in lizards (Sundayet al.,2011; Sundayet al.,2012).In contrast,CTminis lower at high latitudes,likely because species inhabiting lower latitudinal or warmer climatic regions do not face the challenges of cold and seasonally varying temperatures (Janzen,1967;Ghalamboret al.,2006).Although we studied the thermal biology traits ofTakydromuslizards across a wide latitude range,our results are preliminary and can be improved upon by incorporating more species and populations.
The resting metabolic rates ofT.amurensiswere thermally dependent,with allometric enhancement as the temperature increased from 18 °C to 38 °C (Figure 2b).As the basic biological rate,the resting metabolic rates of lizards vary among species,populations,and individuals.The resting metabolic rates ofT.amurensiswere lower than those of the desert-dwelling lizardsE.argus,E.multiocellata,andP.przewalskiiat even temperatures,although they are distributed at similar latitudes (Liet al.,2017; Wanget al.,2019).Unfortunately,the resting metabolic rate ofT.septentrionalis,T.wolteri,andT.sexlineatushas not been studied extensively; this means we were unable to make comparisons amongTakydromuslizards.The sprint speeds ofT.amurensisand otherTakydromusspecies were enhanced as the temperatures increased until the optimal temperature was reached,and then decreased as the temperature increased beyond the optimal temperature (Jiet al.,1996a; Zhang and Ji,2004).However,in all four species we compared,T.amurensishad the lowest sprint speed (Figure 4a).Notably,the optimal body temperature for the sprint speed ofT.amurensiswas 33.20 °C.In the other three species,the optimal temperatures for sprint speed increased as the latitude decreased (Figure 4b).For species from warm and stable thermal environments,sprint speed is more thermally dependent,and the optimal temperatures are higher,as shown inT.sexlineatus(Figure 4; Zhang and Ji,2004).WhyT.amurensis,a cold-climate species,has such a high optimal temperature for sprint speed is still unclear.However,the optimal temperature for sprint speed (i.e.,33.20 ℃),which is roughly equivalent to Tsel(i.e.,32.55℃),imposed an optimal/suboptimal sprint speed at TselinT.amurensis.Future studies could investigate the effect of temperature at a finer scale (e.g.,2 °C between each test temperature),to further clarify the thermal dependence of sprint speed inT.amurensisand thus determine the optimal temperature more precisely.Although the mechanism is unclear,Tseland Toptmight be subject to both thermal environment and confounding evolutionary factors (Huey and Kingsolver,1989).Alternatively,maintaining Tbat a level that appears suboptimal may allow the animal to remain responsive to environmental variation and,if required,increase activity such as those associated with foraging and escaping,following the‘suboptimal is optimal’ hypothesis (Martin and Huey,2008).For example,the range of Teand Tbfor some Liolaemidae lizards is below both their preferred body temperature and the optimal temperature at which they can reach maximum locomotor performance (Boninoet al.,2011).Addtionally,Tbat suboptimal temperatures is also beneficial to avoid over-heating effects in performance,since the performance would drop sharply if the optimal temperatures are surpassed (e.g.,Martin and Huey,2008).
Based on our current preliminary pattern of Tsel,CTmin,CTmax,and Toptfor sprint speed inTakydromuslizards,it is plausible thatT.amurensiswould be less vulnerable to the warming temperatures imposed by climate change than otherTakydromusspecies in habitat in low latitudes.First,the CTmaxforT.amurensisis high; thus,this may indicate that the thermal safety margin is large.However,we were unable to verify this as we did not calculate the thermal safety margin owing to the lack of precise data on environmental temperatures (Sundayet al.,2014).Second,asT.amurensisinhabit areas with cooler temperatures than their physiological optima,it is possible that warming temperatures may enhance their fitness,as in high-latitude species (e.g.,Cabezas Carteset al.,2019;Gómez Aléset al.,2019).
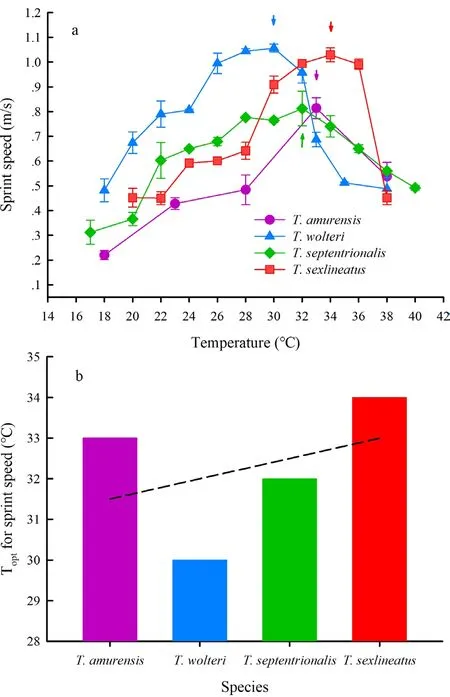
Figure 4 Sprint speed of Takydromus lizards (a) and their Topt (b).(a) Arrows indicate the Topt for sprint speed,respectively.Data are expressed as mean ± SEM.Species are listed from high to low latitude in X axis.Data are collected by Plot Digitizer (http://plotdigitizer.sourceforge.net/),from the figures of references (Ji et al.,1996a; Xu and Ji.,2006; Zhang and Ji,2004).
The thermal biology traits ofT.amurensiswere collected during the reproductive season (i.e.,late April).However,it is possible that seasonal plasticity or acclimatization/acclimation can influence thermal biology traits and their associated performance curve in reptiles and thus cause variations of thermal biology traits inTakydromuslizards.For example,Takydromus septentrionalisfrom different locations did not differ in their selected body temperatures and thermal tolerance after identical thermal acclimation(Yanget al.,2008).Furthermore,the thermal performance curve of the sprint speed ofPlestiodon chinensiswas shown to vary between seasons (Sunet al.,2014).The metabolic enzyme activity in the muscles ofAlligator mississippiensisalso responds to temperatures differently between seasonal acclimation (Seebacheret al.,2003).Determining the seasonal variation of biological traits in cold-climate species such asT.amurensisis important because thermal fluctuations are predicted to be more significant at high latitudes (IPCC 2013).Another potential cause of the observed variation in the thermal biology traits ofT.amurensisin the current study may be the limited sample size (i.e.,n=13).Larger sample sizes of individuals will be required to investigate seasonal acclimation and confirm our preliminary results on latitudinal trends.However,as a cold-climate species,the natural population density ofT.amurensisis low and is threatened by habitat destruction (Zhao,2002; Portniaginaet al.,2019).In future studies,meta-analysis methods may be more suitable for revealing the geographical patterns of thermal biology traits within congeners.It is possible that the inter-specific variation in thermal biology traits along latitudinal gradients is attributed to other factors,such as precipitation,vegetation cover,or food availability,which also varies across altitudes.However,as theTakydromusspecies occupy similar elevations,it is plausible that the observed pattern among species is induced by latitudinal differences.Future studies,investigating more species distributed over a large geographical span,should be conducted to test altitudinal pattern.
Our study determined the thermal biology traits of cold-climate distributedT.amurensis,summarized the interspecies variation,and provides preliminary findings suggesting that Tsel,CTmax,and CTminvary along latitudinal gradients inTakydromuslizards.These findings will contribute to the understanding of the thermal adaptation of reptiles and establish basic criteria for determining the climatic vulnerability ofTakydromusspecies.Future studies should investigate more details of the biological response to thermal environments,including acclimation and life history variations.Furthermore,building on the data collected in this study with additional measurements of operative temperature and active body temperature in field,we will be able to calculate the thermal safety margin and the effectiveness of thermoregulation,which would be helpful in evaluating the degree of safety or beharioral flexibility to thermal variation,in the context of climate warming.Additional research into thermal biological responses will facilitate projection of the vulnerabilities ofTakydromusspecies to climate warming.
AcknowledgementsWe thank Xingzhi HAN,Teng LI and Tingting WANG for their assistance in the field and Lab.Ethics approval and protocol (IOZ14001) for the collection,handling,and husbandry of the study animals was given by Animal Ethics Committees at Institute of Zoology,Chinese Academy of Sciences.This work was supported by National Natural Science Foundation of China (31870391 and 31500324).Baojun SUN is supported by Youth Innovation Promotion Association CAS (No.2019085).
 Asian Herpetological Research2020年4期
Asian Herpetological Research2020年4期
- Asian Herpetological Research的其它文章
- Characterization of Skin Symbiotic Bacteria of Sympatric Amphibians in Southeastern China
- The Role of Environmental Stress in Determining Gut Microbiome:Case Study of Two Sympatric Toad-headed Lizards
- Adaptive Evolution of the Ventral Scale Microornamentations among Three Snake Species
- Strong Limb Tactics of the Boulenger’s Lazy Toad,Scutiger boulengeri:Inferred from Limb Muscles
- The Relationship between Shell Morphology and Crevice Size Affecting Retreat Selection of the Keeled Box Turtle (Cuora mouhotii)
- Spring Voices in Korean Rice Fields:The Effect of Abiotic Variables and Syntopic Calls on the Calling Activity of the Treefrog Dryophytes suweonensis