
ATP gatekeeper of Plasmodium protein kinase may provide the opportunity to develop selective antimalarial drugs with multiple targets
2020-07-20 02:45:20FauzeMahmudPingChinLeeHabibahWahabKhairulMohdFadzliMustaffaChiuanHerngLeowAzharRasulNgitShinLai
Fauze Mahmud, Ping Chin Lee, Habibah A Wahab, Khairul Mohd Fadzli Mustaffa, Chiuan Herng Leow,Azhar Rasul, Ngit Shin Lai?
1Institute for Research in Molecular Medicine, Universiti Sains Malaysia, Penang, Malaysia
2Faculty of Science and Natural Resources, Universiti Malaysia Sabah, Sabah, Malaysia
3Faculty of Pharmaceutical Science, Universiti Sains Malaysia, Penang, Malaysia
4Department of Zoology, Government College University Faisalabad, Pakistan
ABSTRACT Malaria is one of the most devastating infectious diseases that caused millions of clinical cases annually despite decades of prevention efforts. Recent cases of Plasmodium falciparum resistance against the only remaining class of effective antimalarial(artemisinin) in South East Asia may soon pose a signi ficant threat.Hence, the identi fication of new antimalarial compounds with a novel mode of action is necessary to curb this problem. Protein kinase has been implicated as a valid target for drug development in diseases such as cancer and diabetes in humans. A similar approach is now recognized for the treatment of protozoan-related disease including malaria. Few Plasmodium protein kinases that are not only crucial for their survival but also have unique structural features have been identi fied as a potential target for drug development. In this review, studies on antimalarial drug development exploiting the size of Plasmodium protein kinase ATP gatekeeper over the past 15 years are mainly discussed. The ATP-binding site of Plasmodium protein kinases such as Pf CDPK1, Pf CDPK4, Pf PKG, Pf PK7,and Pf PI4K showed great potential for selective and multi-target inhibitions owing to their smaller or unique ATP-gatekeeper amino acid subunits compared to that of human protein kinase. Hence it is a feasible solution to identify a new class of active antimalarial agents with a novel mode of action and longer clinical life-span.
KEYWORDS: Plasmodium falciparum; Protein kinase inhibitor;ATP-binding site; Antimalarial activity
1. Introduction
Since its first description in ancient Egypt texts, malaria remains one of the most life-threatening and widespread infectious diseases in the world. Currently, sixPlasmodiumspecies [Plasmodium (P.)falciparum, P. vivax, P. malariae, P. ovale curtisi, P. ovale wallikeri and P. knowlesi] are known to cause malaria in human, of whichP.falciparumis the most virulent[1]. This species of malarial parasite caused almost half a million deaths annually, especially in the sub-Saharan region, while mild but prevalent malaria cases in Asia and South America are caused byP. vivax. The malarial protozoan is transmitted to humans by femaleAnophelessp. mosquitoes, and its pathological symptoms start within one week to months after infection, depending on thePlasmodiumspecies[2]. A global effort to eliminate malaria using both artemisinin-based combination therapy (ACT) and insecticide-treated bed nets since the 1950s has signi ficantly reduced the malaria mortality rate[3,4].
Although the infection rate has steadily been decreasing by 48%in total over the past decade, malaria still represents a signi ficant human and economic burden, with more than 1 billion malaria cases recorded worldwide from 2001-2015[3,4]. The infection continues to put 3.2 billion people, especially children under the age of five,at risk as recent data indicate that 216 million malaria cases were reported in 2016, increasing by 5 million cases compared with 2015[3,4]. Currently, only four classes of antimalarial compounds available in clinical use which comprises of artemisinin derivatives,quinine or other aminoquinolines, antifolates, and hydroxyl naphthoquinone atovaquone[5]. The recent con firmed development of artemisinin resistance in South East Asia further complicates this problem[6,7]. As we are on the verge of yet another signi ficant development ofPlasmodiumresistance strain since the emergence of chloroquine-resistance, the effort of finding new antimalarial drugs,especially with a novel mode of action or target has now become more critical than ever.
As in 2018, a total of 13 new lines of antimalarial drugs are now tested at phaseⅠand beyond, including tafenoquine (analog of primaquine), which has been recently approved by US Food and Drug Administration (FDA)[8]. Commercialized as Krintafel,tafenoquine is used for the treatment of malaria caused byP. vivaxduring both blood and tissue stage besides acting as gametocytocidal.Other multi-stage acting drugs in the clinical phases include KAF156(PhaseⅡ)[9,10], and M5717 (PhaseⅠ)[11]. Despite their potential, the mechanisms of action of both KAF156 and M5717 are unknown[12].Without the knowledge of the exact target, hit progression will become more challenging[13].
The use of antimalarial drugs with the unknown target is nothing new. Most antimalarial drugs in the market were developed without the knowledge of known drug target as their therapeutic potency have only been identi fiedviacell-based assay[14]. Determining the exact target of a drug is valuable knowledge as it can prevent latestage failures and increase the chances of drug approval. Information on the actual target also leads to better dosing, helps to monitor potential side effects of a drug, and stratify better clinical trials on suitable patients[15]. Thus, experimental design targeting selected targets such as protein or lipid, is an appropriate approach to solve this issue[16].
For this reason, MMV390048 is one of the most exciting new antimalarial drug lines as it is the firstP. falciparumprotein kinase (PfPK) inhibitor reaching clinical validation, targetingPfPhosphatidylinositol 4-kinase (PfPI4K). This drug is acting as blood schizonticide that inhibits gametogenesis and oocyst formation[17].BesidesPfPI4K, the onlyPfPK validated clinically, few other protein kinase targets are now in development as potential drug targets such asPfCDPKs (PfCDPK1,PfCDPK4),PfPKG andPfnek-1 (all genetically, phenotypically andin vivovalidated),PfMRK,PfGSK-3, andPfPI3K (all genetically and phenotypically validated),PfFIKK8 (phenotypically validated) andPfPKA(genetically validated) have also been identi fied[18].
All these protein kinases are veri fied to be essential forPlasmodiumsurvival, and most of them are expressed at different stages ofPlasmodium’s life cycle with varying importance. Infected mosquitos transfer the sporozoites into the bloodstream, and it travels to the liver (PfPK7 andPfPI4K are essential at this stage) (1)[19]. In the infected hepatocytes cells, sporozoites then matured into schizont,and finally, the merozoites released into the bloodstream and infect red blood cells for the asexual stage (essentialPlasmodiumprotein kinases arePfCDPK4 andPfCDPK6) (2)[20,21]. The merozoites then developed into ring-stage/early trophozoite, late/mature trophozoite, red blood cell (RBC) schizont, and finally merozoites are released to infect more RBC. At this stage, manyPfPK were identi fied to be involved such asPfPI4K,PfFIKK8,PfCDPK4, andPfGSK-3 (3)[22,23]. Instead of progressing into RBC schizont, some of the trophozoites will develop into male and female gametocytes that will be taken up by the mosquito for the sexual stage, and the cycle is repeated (4) (Figure 1).
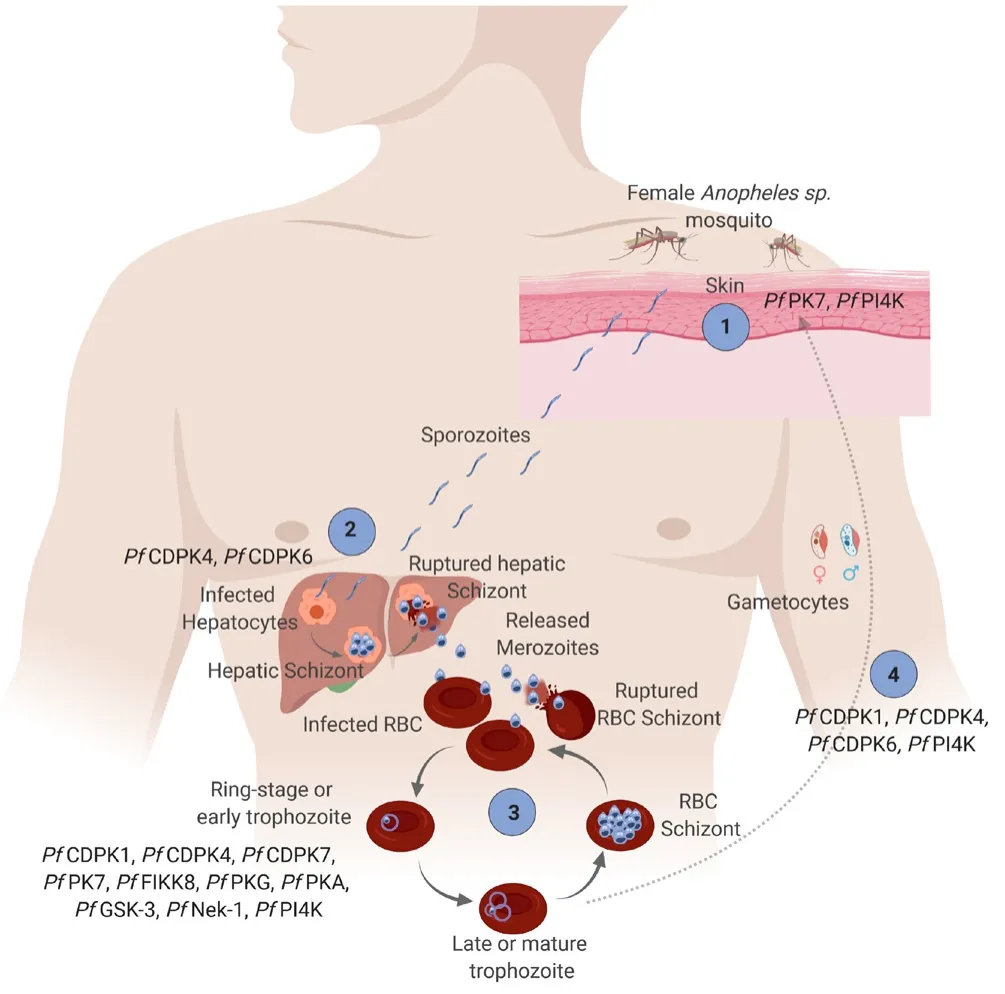
Figure 1. Stage-speci fic involvement of potential Plasmodium falciparum kinase at different stages of its life cycle. (Figure was drawn using BioRender online software).
PfPK is also an emerging target for antimalarial drug development due to the success of protein kinase inhibitors in humans in which the US FDA has approved 33 human protein kinase inhibitors to date. Almost 100 eukaryotic protein kinase-related (ePK-related)enzymes have been identi fied inP. falciparumkinome, where most of them can be grouped into seven ePK families. Due to the long independent evolution ofPlasmodiumkinome, somePfPK cannot be categorized into any ePK such asPfPK6 andPfPK7 and known as “orphan kinase”. Moreover, some of these orphan kinases have unique features with no orthologue in humans, such asPfCDPKs(higher resemblance to plant protein kinase) andPfFIKKs (can only be found in the phylum of Apicomplexa)[18,24].
Furthermore, recombinantPfPKs are adaptable for high-throughput screening (HTS) format in which a screening using thousands of compounds has become a powerful and robust tool to identify potential new antimalarial compounds. One of the largest and most diverse compound libraries screening performed to date utilized 1.7 million compounds in which fully integrated and automated high-throughput fluorescence-based phenotypic assay (1536-well format) was applied. This study identified approximately 6000 small molecules (with more than 500 distinct scaffolds) that showed potent antimalarial activity (<1.25 μM). Over 95% of the identi fied active scaffolds were not previously reported as having antimalarial activity[25].
Another study was performed in which almost 2 million compounds in GlaxoSmithKline’s library were screened againstP. falciparum3D7 and Dd2 culture in 384-well. The antimalarial potential was measured based on the LDH activity, of which almost 14000 compounds were con firmed with more than 70% parasite clearance at 2 μM concentration. Also, more than 8000 compounds showed potent activity against the multidrug-resistant strain Dd2. More than 80% of the identi fied active compounds were never previously described with antimalarial activity[25,26].
Both studies postulated thatPfPK might be the possible target based on the mechanism of action (MOA) historical data or structure-activity relationship (SAR) analysis of compounds with known activity. For instance, SAR analysis revealed that out of 6000 active compounds, 48 active ligands sharing the same scaffold as staurosporine, a well-known protein kinase inhibitor[22]. Meanwhile,MOA analysis indicated that almost 50% of the active compounds are targeting protein kinase. At this point,PfPKs are strongly suggested as a potential yet unexploited antimalarial drug target as its inhibitor not only could potentially inhibit multiplePfPKs but also distantly related enzymes due to structural similarities of the binding sites. However, this may also pose a risk to humans as it might allow the inhibition of related human protein kinases[23].
As a follow up to the HTS cell-based assay, the first step typically is to identify the actual protein kinase targeted by the inhibitor that caused antimalarial activity observed in the phenotypic assay. Next,is to identify druggablePfPK that has unique structural features for speci fic inhibition over human protein kinase[18]. Thus, the critical structural difference betweenPfand human protein kinases could be identi fied to minimize speci ficity concern due to interspecies structural similarities. ATP- and substrate binding sites of the protein kinase are usually the primary target for ligand binding as they directly control protein kinase activation and activity. Some studies reported the differences between human andPlasmodiumprotein kinase, especially on the ATP-binding pocket of glycogen synthase kinase-3 (GSK-3). It was revealed that subtle structural differences which may provide a certain degree of speci ficPlasmodiumprotein kinase inhibition[27-29].
2. ATP-binding site of P. falciparum protein kinase provides a potential target for drug development
Sequencing of human/Homo sapiensGSK-3 (HsGSK-3) andP. falciparumGSK-3 (PfGSK-3) revealed low conservation in both domain identity and similarity, 56% and 76% similarities,respectively. Computational analysis ofPfGSK-3 withHsGSK-3 using SYBYL further revealed that the ATP-biding site ofPfGSK-3 is slightly smaller and less extensive compared withHsGSK-3β.Also, the hydrophobic sub-pocket at the bottom of thePfGSK-3 ATP binding site is protected by Met137 (Leu135 inHsGSK-3).Four out of 11 probes were shown to have different binding strength inPf- andHsGSK-3 ATP-binding sites. OC1 probe (H-bond acceptor probe) is shifted towards the inner region of thePfGSK-3 ATP-binding site as no salt bridge formed between Gln162 and Lys166 to arrest its con firmation (formed in human)[27].
N2 (H-bond donor probe) interact with both carbonyl groups inPf-/HsGSK-3. However, signi ficant molecular interaction fields (MIFs)difference still can be observed due to differences in nearby residues.N1 probe has stronger MIFs forPfGSK-3 as its binding is affected by the backbone carbonyl of Ala106/Ala83 residues near gatekeeper at a distance of 3.34 ? (3.17 ? in human). Iodine (I) (van der Waals radius of 2.15 ?) shows strong interaction withPfGSK-3 as it faced less steric hindrance at the bottom ofPfGSK-3 ATP-binding site due to a greater distance of MIF to the sulfur of Cys224/Cys199 (Pf-/HsGSK-3) inPfGSK-3 (4.2 ?) compared withHsGSK-3 (2.4 ?).Hence, this study managed to demonstrate the effect of subunit differences betweenPf- andHsGSK-3 on the binding of a possible inhibitor[27].
Another bioinformatics study on human and parasite GSK-3 managed to demonstrate inhibitor selectivity differences due to the size of the ATP-gatekeeper[28]. The activity of paullone (ATP-competitive inhibitor) and its derivatives were found to have a higher affinity towardsHsGSK-3 by 30 to 300-fold compared withPfGSK-3. As previously indicated, it is likely caused by methionine gatekeeper inPfGSK-3 that is longer and more flexible than the leucine subunit ofHsGSK-3[30]. It created a steric effect on paullone and its derivatives. These amino acids are termed as“ATP gatekeeper” because they guard the access to the unexploited hydrophobic pocket[31].
In addition, 3,6-diamino-4-(2-halophenyl)-2-benzoylthienol[2,3-b]pyridine-5-carbonitriles (5v) was identified as a new class ofPfGSK-3 inhibitor. Its derivatives with 2-ClPhe and 3-ClPh on R1and R2respectively showed the most potent activity (PfIC50is 5.5 μM).The IC50of 5v onPfGSK-3,HsGSK-3α, andHsGSK-3β is 0.48 μM,>100 μM, and 3.0 μM, respectively.In silicostudy showed that better inhibition ofPfGSK-3 achieved by 5v was due to hydrophobic interaction between the thiophene of 5v with ATP-gatekeeper subunit ofPfGSK-3 (methionine). This interaction was not formed inHsGSK-3 as its gatekeeper subunit (Leucine) is located further[29].Hence, the size ofPfPK ATP-gatekeeper is a compelling feature to be exploited for drug discovery as it can control which inhibitor to bind in the ATP-binding site[32].
Currently, not manyPfPKs ATP binding site has been described.However, most of thePfPKs have small or very small ATP gatekeepers, as approximately 30PfPKs (of 86 to 99 knownPfPks) have threonine or serine ATP gatekeeper subunit[21,33]. Besides that, somePfPKs possessing bulky yet with unique amino acid subunits (tyrosine) that is very rare in humans[18,34-36]. In contrast,human protein kinase usually has bulky gatekeeper residues such as methionine, leucine, and phenylalanine. Only 19% of human protein kinase has a small ATP gatekeeper (threonine)[37,38]. This structural difference is significant as a bulky gatekeeper blocks the hydrophobic pocket located behind ATP-binding sites, and a small gatekeeper exposes this pocket to inhibitors[36]. Thus, it can be exploited for the identi fication of speci fic inhibitors termed as“bumped kinase inhibitor” that relatively has low off-target effects on the human kinome[34,39].
To targetPfPK ATP-binding sites, scaffolds such as purine that are structurally related to ATP (natural substrate of all protein kinase) were indicated as a potential inhibitor[27]. Purfalcamine(2,6,9-trisubstituted purine) was identi fied as aPfCDPK1 inhibitor that causedP. falciparumto accumulate in the late schizogony and failed to egress from the merozoite stage. In the ATP-binding site ofPfCDPK1, nonconserved residues such as Arg60, Glu149,and Lys202 can be targeted to establish a salt bridge for better selectivity. Moreover,PfCDPK1 has small-sized ATP gatekeeper residue (threonine) protecting the bottom of the ATP-binding site.Interestingly, purfalcamine was also indicated to inhibitPfCDPK5(highest homology withPfCDPK1, but with bulky ATP gatekeeper)with lower affinity (IC50of 17 nm inPfCDPK1 and 3.5 μM inPfCDPK5, respectively). Further test on four mammalian cell lines in which the EC50(230 nm) suggested the therapeutic window of purfalcamine was 23- to 26- fold[40].
A similar strategy of using a pyridine motif and another aromatic linker such as pyrimidine (class 1) and fluoropyridine (class 2) attached to imidazopyridazine was recently applied. Class 1 compound was found to be active against bothPfCDPK1 andPfPKG. The ATP-binding site ofPfCDPK1 andPfPKG is closely related and shared the same sequence homology that includes gatekeeper residues, threonine at Thr145 (PfCDPK1) and Thr618(PfPKG), which may explain dual inhibition exerted by class 1 inhibitor. The substitution ofPfCDPK1 gatekeeper with bulky residue such as glutamine signi ficantly reduced the sensitivity ofPfCDPK1 to these inhibitors as it blocks the access to the ATP-binding site. A series of class 1 inhibitors were tested in which most of them showed a higher affinity towardsPfPKG, notably class 1-compound A (IC50PfPKG: 0.002 μM and IC50PfCDPK1: 0.008 μM,respectively). This compound also almost 6000 more potent on wild typePfPKG than mutant protein (PfPKG with glutamine ATP-gatekeeper)[38].
A recent study on the potential ofPfPKG as a druggable protein was explored using the imidazopyridine series. Imidazopyridine(with the addition of cyclopropylmethylene group) (ML10) was identi fied as the most potent compound (160 pM). ML10 caused merozoite failed to egress and block transmission ofP. falciparumgametocytes toAnophelessp. Until today,PfPKG crystallization with or without inhibitor is still unable to be achieved. Thus, the interaction between ML10 on thePlasmodiumATP-binding site was studied based on the co-crystal structure ofP. vivaxPKG (PvPKG).Amino-pyrimidine of ML10 formed a hydrogen bond with the backbone of Val614 (Val621 inPfPKG), mimicking ATP. Also, the sulfonamide group of ML10 formed a hydrogen bond with Asp675(Asp682 inPfPKG) and Phe676 (Phe683 inPfPKG) of the DFG triad for stronger binding. Remarkably, the fluorophenyl group can interact with the hydrophobic pocket ofPvPKG that is guarded by threonine (Thr611 inPvPKG and Thr618 inPfPKG), a small gatekeeper[39,41].
Moreover,PfPKG has been recognized to have distinct properties than human PKG[42]. Hence, such interaction is blocked by a large subunit that making human PKG insensitive against ML10.Interestingly, ML10 also showed very little inhibitory activity against 80 human protein kinase panels representing all kinase families including human kinase protein with small ATP gatekeeper(tested at 100 μM)[41]. Besides,PfCDPK1 was found dispensable for the survival ofPlasmodiumduring the red blood stage. Instead,the antimalarial activity of class 1 was found due to the inhibition ofPfPKG during red blood stage. AlthoughPfCDPK1 was suggested as not a suitable target for blood schizonticide development, it is the potential target for antimalarial acting during the gametocytes stage[43,44]. Hence, a compound such as class 1-compound A may provide selective antimalarial that can act on different stages ofPlasmodiumlife-cycle.
PfPKs with bulky but unique ATP gatekeepers notably,PfPK7 andPfPI4K, also showed potential as drug targets. A series of pyrimidine and pyridazine class of compounds were identi fied to inhibit the activity ofPfPK7, in which the most potent compound is (S)-4-[6-(1-hydroxy-3-methylbutan-2-ylamino)imidazo[1,2-b]pyridazin-3-yl] benzonitrile (K510). K510 was shown to mimic ATP by forming a hydrogen bond with Met120 (backbone amide group), and heterocyclic ring established interactions with Leu34 and Leu179. The interaction is further strengthened by the dipolar interaction between K510 nitrile moiety and the hydroxyl group ofPfPK7 gatekeeper (Tyr117)[34]. Another study described imidazopyridazine derivative-34 with antimalarial activity (Pf3D7 IC50of 1.03 μM) and a selectivity index of 23.32. Although the binding site of derivative-34 is unknown, the replacement of larger substituent (amine, in derivative-22 to 24), resulted in signi ficant antimalarial activity decrease[45].
Speci fic inhibition ofPfPK despite having a bulky ATP gatekeeper subunit was confirmed achievable, using imidazopyrazines and quinoxalines scaffolds againstPfPI4K. Imidazopyrazines KDU691 and KAI407 were identi fied to targetPfPI4K ATP-binding site, but both displayed excellent selectivity over human lipid and protein kinases.In silicoanalysis revealed that the N1 of KAI407 imidazole ring formed a hydrogen bond mimicking hydrogen bonds made by the adenine of ATP[18,46,47]. The activity of these compounds on PI4K was further confirmed based on enzymatic assay against recombinantPvPI4K. Their inhibitory activities are ATP concentration-dependent, which indicates the ATP-competitive mode of action. Also, KA1407 showed no effect on a panel of human protein kinases consisting ofHsPI4KIIIα andHsPI4KIIIβ[46].
Recently, aminopyridine was identi fied as another potential class of compounds that act as potent and selectivePfPI4K inhibitor resulting in the identi fication of 2-aminopyridine MMV390048, the firstPfPK inhibitor reaching the clinical stage. This compound was recognized to act as an ATP-competitive inhibitor, as well[48]. Its efficacy targetingPfPI4K was described in-depth from the screening of a 2-aminopyridine class of small molecule. MMV390048 potency was de fined based on a humanized mouse model, mouse-to-mouse transmission, and monkeys. This inhibitor is indicated to be active against allPlasmodiumlife-cycles except for late hypnozoites in the liver.PfPI4K was con firmed as its molecular target from genomic and chemoproteomic studies. Interestingly, the kinobeads analysis revealed that MMV390048 was shown to binds only on the ATP-binding site ofPfPI4K but not to its human analog (both PI4Kα and PI4Kβ) indicating selective inhibition[17].Based on these findings, the ATP gatekeeper ofPfPK is a potential structural feature to be exploited for the development of antimalarial agents as it provides selective inhibition by specific chemical scaffolds towardsPlasmodiumprotein kinase that reduce unwanted risk in human[49].PfPKs with small gatekeeper was also shown to be inhibited by the same active compound such asPfCDPK1 andPfPKG both inhibited by imidazopyridazine, and compound BKI 1294 inhibitedPfCDPK1 andPfCDPK4 (Table 1)[33,44].Interestingly, somePfPKs with small and bulky gatekeepers (PfPK7,PfPKG, andPfCDPK1) were also reported as the targets of imidazopyridazines derivatives, owing to the overall similarities of their ATP-binding sites[44,45,52]. A similar finding was reported in which various scaffolds able to inhibitPfCDPK1,PfCDPK4,PfPK6, andPfPK7 at the same time[53].

Table 1. List of inhibitors targeting the ATP-binding site of Pf PK.

Table 1. Continued.
Besides, the exploration ofPfPKs with very small ATP gatekeepers(especially thePfFIKK family) might shed more light on their potential to develop a new line of antimalarial agents[18,24]. In all,protein kinase inhibitor targeting the ATP-binding site ofPfPKs may not only work well for speci fic inhibition but also with multiple targets inPlasmodiumas well, partly contributed by the size ofPfPKs ATP gatekeepers that are divergent than human protein kinase[54-57].
3. Conclusions
The size of the ATP-gatekeeper has been indicated as potential structural features to provide specificPlasmodiuminhibition as they are generally smaller than that of human protein kinase. The main advantage of this approach is the selective inhibition ofPfPKs over human protein kinase. ATP-binding may also lead to the identi fication of antimalarial drugs with multiple targets asPfPKs generally conserved among them. Hence, it may eventually delay the development of drug resistance strain, an assumption based on slow resistance development against artemisinin (almost 40 years), that is believed to act on multiple targets as well (all stages of malaria)[58,59]. Overall, a compound that showed a high affinity towardsPlasmodiumthat is in fluenced by the size of itsPfPKs ATP gatekeeper is probably one of the best options for the development of the next antimalarial agent.
Conflict of interest statement
The authors declare that there are no con flicts of interest.
Authors’ contributions
F.M. conceived the idea, drafting, and editing the presented article.L.P.C, H.A.W, K.M.F.M, L.C.H, and A.R. were involved in the critical revision of the article. L.N.S is the project leader, contributed to the focus and critical revision of the manuscript.
 Asian Pacific Journal of Tropical Medicine2020年8期
Asian Pacific Journal of Tropical Medicine2020年8期
- Asian Pacific Journal of Tropical Medicine的其它文章
- First COVID-19 related death in Pakistan in a patient with a travel history in Saudi Arabia
- COVID-19 pandemic in Rwanda: An overview of prevention strategies
- Using twitter and web news mining to predict COVID-19 outbreak
- Effectiveness of intermittent preventive treatment in pregnancy with sulfadoxinepyrimethamine: An in silico pharmacological model
- Diagnostic performance of C-reactive protein level and its role as a potential biomarker of severe dengue in adults
- Chinese Expert Consensus on Early Prevention and Intervention of Sepsis