
Signal transducer and activator of transcription 3 promotes the Warburg effect possibly by inducing pyruvate kinase M2 phosphorylation in liver precancerous lesions
2019-05-08 07:18:46YangHuiBiWenQiHanRuoFeiLiYunJiaoWangZunShuDuXueJiangWangYingJiang
World Journal of Gastroenterology 2019年16期
Yang-Hui Bi,Wen-Qi Han,Ruo-Fei Li,Yun-Jiao Wang,Zun-Shu Du,Xue-Jiang Wang,Ying Jiang
Abstract
Key words: Warburg effect;Hepatic progenitor cell;Signal transducer and activator of transcription 3;Pyruvate kinase M2;Liver precancerous lesion
INTRODUCTION
Cancerous cells transform significant amounts of glucose into lactate irrespective of oxygen availability.This phenomenon,also known as aerobic glycolysis or the Warburg effect,meets the energetic demands of tumor cells and provides them with growth and survival advantages.The Warburg effect is frequently enhanced in hepatocellular carcinoma (HCC)[1,2].Many studies have shown that HCC is closely associated with precancerous lesions.Liver precancerous lesions are well-established histopathologically premalignant tissue with the highest risk for tumorigenesis and are critical for hepatocarcinogenesis in human and rodents[3-5].Recognition and treatment at the precancerous stage might reduce the incidence and mortality of HCC.However,the underlying molecular mechanism for the development of liver precancerous lesions remains unclear.
The liver stem cell differentiation arrest theory and the dedifferentiation of mature hepatocytes theory are two widely accepted hypotheses of hepatocarcinogenesis.Studies have noted a progenitor cell phenotype in many HCCs.Alterations in the hepatic niche can lead to the malignant transformation of hepatic progenitor cells[6-8].Signal transducer and activator of transcription 3 (STAT3) is a member of the STAT family of transcription factors and is activated in several cancers with a poor prognosis,such as HCC,breast cancer,and esophageal squamous cell carcinoma[9-11].Once it is activated by tyrosine phosphorylation,STAT3 translocates into the nucleus and regulates the expression of target genes,mediating numerous aspects of tumorigenesis,including proliferation,angiogenesis,immunity,motility,apoptosis,and cell differentiation[12,13].In addition,STAT3 can regulate aerobic glycolysis in breast cancer cells by stimulating the transcription of hexokinase 2[14].STAT3 also upregulates lactate dehydrogenase A and pyruvate dehydrogenase kinase 1 to increase glycolysis in myeloma cells[15].However,whether activation of STAT3 enhances the Warburg effect in liver precancerous lesions and the underlying mechanism remain unclear.
Pyruvate kinase (PK),a key enzyme in glycolysis,catalyzes the synthesis of pyruvate and ATP,using phosphoenolpyruvate (PEP) and ADP as substrates.There are four mammalian pyruvate kinase isoforms:PKL,PKR,PKM1,and PKM2.PKL is mainly expressed in the liver and kidney,and PKR is highly expressed in erythrocytes.PKM1 is preferentially expressed in adult tissues,while PKM2 is abundantly expressed in rapidly proliferating tissues,embryos,and tumors[16-18].Recent studies have shown that PKM1 can promote tumor cell proliferation and activate glucose catabolism in small-cell lung cancer (SCLC)[19].But more results confirm that among the four mammalian pyruvate kinase isoforms,PKM2 is most closely related to tumor cell proliferation.PKM2 is an important regulator of the Warburg effect and is highly expressed in many tumors.Tetrameric PKM2 has PK activity in normal cells.PKM2 tetramers are tyrosine-phosphorylated at Y105 to transform dimeric PKM2 in tumor cells[20,21].Then,PKM2 dimers promote the synthesis of deoxynucleotides,providing tumor cells with a growth and proliferation advantage.Dimeric PKM2 can enhance aerobic glycolysis,increase lactate production,and regulate the expression of genes that mediate the Warburg effect in many tumors[18,20],including HCC[22].However,whether STAT3 activation enhances the phosphorylation of PKM2 in cancer is unknown.
Here,we showed that Warburg effect was initiated in liver precancerous lesions in rats and STAT3 phosphorylation was involved in liver precancerous lesions probably by activating hepatic progenitor cells in the Solt-Farber model.Furthermore,we confirm that STAT3 activation promoted the Warburg effect by upregulating p-PKM2 in transformed hepatic progenitor cells in rats.Our study can help better understand the role of STAT3 and PKM2 in HCC formation.
MATERIALS AND METHODS
Experimental animals
Sixty adult male Wistar rats (6 wk old),weighing 180-200 g,were purchased from Beijing Vital River Laboratory Animal Technology Co.Ltd (Beijing,China).All animal feeding and experimental procedures were approved by the Ethical Committee of Capital Medical University and were performed according to National Institutes of Health (NIH) Guidelines.A modified Solt-Farber model of liver precancerous lesions was generated as described[23,24].The animals were acclimatized to laboratory conditions (23 °C,12 h/12 h light/dark,50% humidity,ad libitumaccess to food and water) for 1 wk prior to experimentation.Then,a single dose of diethylnitrosamine(DEN,200 mg/kg body weight;Sigma,St.Louis,MO,United States) was injected intraperitoneally.After a two-week recovery,the rats were subcutaneously inserted with 2-acetylaminofluorene (2-AAF;Innovative Research,Miami,FL,United States)pellets (50 mg/pellet over a 21-d release) for 1 wk followed by a two-thirds partial hepatectomy (PH).The animals were then euthanized nine days after PH.The livers of the rats were removed rapidly and dissected.Rats with no treatment,rats exposed to DEN and PH,and rats exposed to AAF and PH were used as controls.
Cell culture
The WB-F344 rat hepatic progenitor cell line (a gift from Dr.Geng-Tao Liu,Peking Union Medical College) was cultured in Dulbecco’s Modified Eagle Medium and Nutrient Mixture F-12 (DMEM/F12) (Hyclone) with 100 U/mL penicillin and 100 μg/mL streptomycin with or without 10% fetal bovine serum (FBS;Corning,KS,United States).The cells were maintained in the logarithmic growth phase at 37 °C in 5% CO2.We induced the malignant transformation of WB-F344 cells according to a previous study[25,26].Briefly,WB-F344 cells were exposed to 3 μg/mL N-methyl-N’-nitro-N-nitrosoguanidine (MNNG) for 24 h,and then the cells were treated with 7 ×10-7mol/L H2O2for 12 h per day for 21 d.WB-F344 cells with no treatment was cultured as controls.
Histopathology
Rat livers were fixed in formalin for 24 h,paraffin-embedded,and sectioned into 5-μm-thick slices for hematoxylin and eosin (HE) staining.
Immunohistochemistry
Immunohistochemistry was performed as previously described.Sections were incubated with rabbit anti-PKM2 (1:800;CST,MA,United States) and rabbit antiglutathione S-transferase-π (GST-π;1:1000;MBL,Nagoya,Japan) overnight at 4 °C.The appropriate secondary antibody (goat anti-rabbit IgG,ZSGB-BIO,Beijing,China)was applied for 30 min,and 3,3’-diaminobenzidine was used as the chromogen.Negative controls were run for each antibody,using PBS instead of the primary antibody.Representative images were captured with an Axio Imager 2 (Zeiss,OBK,Germany).
Immunofluorescence
Immunofluorescence was conducted as described.The slices were incubated at 4 °C overnight with mouse anti-OV-6 antibody (1:150;Roche,Basel,Switzerland) and rabbit anti-PKM2 antibody (1:50;CST) or rabbit anti-p-STAT3 antibody (1:100;CST),followed by fluorescent staining with FITC-conjugated anti-mouse IgG and Alexa Fluor 594-conjugated anti-rabbit IgG (Jackson,PA,United States).DAPI was used to stain the nuclei in the tissue samples.Images were captured with an Axio Imager 2.
Soft agar assay
Cell were assessed for colony formation in soft agar assays using a Cell Biolabs Cytosolic Cell Transformation Assay kit (Cell Biolabs,CA,United States) as per manufacturer’s instructions.
Analysis of glucose consumption and lactate production
Liver tissue samples were lysed in ice-cold normal saline (0.3%).Cells were seeded in 6-well plates (8.5 × 105cells/well).The glucose and lactate concentrations in the medium and liver tissue homogenate were measured by the glucose-oxidase method(Applygen Technologies,Beijing,China) and with a lactic acid assay kit (Nanjing Jiancheng Biotechnology,Nanjing,China),separately.The glucose consumption and lactate production were normalized to protein concentration and cell numbers.
Cell cycle and aneuploidy cells assay
The cell cycle was examined by flow cytometry.Cells (106cell/mL) were harvested and fixed in 75% alcohol at 4 °C overnight.After a wash step in PBS,500 μL of cell cycle reagent propidium iodide (PI)/RNase staining solution (CST,MA,United States) was added to each tube and incubated for 30 min at room temperature in the dark.DNA content was analyzed on an EPICS@XL flow cytometer (Beckman Coulter,CA,United States).DNA content of diploid cells in WB-F344 cells was used as an internal standard to determine DNA index (DI).Cells with a DI = 0.95-1.05 were defined as diploid,cells with a DI = 1.92-2.04 were defined as tetraploid,and cells with a DI > 2.04 was considered as aneuploid[27].Three independent experiments were performed for each assay condition.
Cellular ATP level
Cells were seeded in 96-well plates (5 × 104cells/well).At the end of the treatments,the cell resuspension solution was collected.ATP content was measured using the CellTiter-Glo2.0 assay kit (Promega,WI,United States) and normalized to cell numbers.Three independent experiments were performed for each assay condition.
Western blot analysis
Liver tissue samples were lysed in ice-cold RIPA buffer that was supplemented with protease inhibitors.Protein concentrations were determined using a bicinchoninic acid protein assay kit (Thermo,MA,United States).The protein extracts from liver tissue (50-100 μg) and cell lysates (40-60 μg) were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore,MA,United States) for Western blot analysis.The membranes were incubated with primary antibodies at 4 °C overnight and then were incubated with the appropriate secondary antibodies at room temperature.The primary antibodies included antibodies against GST-π (MBL,Nagoya,Japan),alphafetoprotein (AFP;Proteintech,Chicago,United States),GAPDH (GXY,Beijing,China),cytokeratin 19 (CK19),PKM2,p-PKM2,STAT3,p-STAT3,proliferating cell nuclear antigen (PCNA),β-tubulin,and β-actin (CST,MA,United States).The protein bands were visualized using enhanced chemiluminescence detection reagents (Applygen,Beijing,China) and scanned on a FlourChem HD2 System (Protein Simple,CA,United States).
Cell transfection with siRNA
Cells were seeded into 6-well plates (1.0 × 105cells/well) on the day before the transfection.The siRNA for PKM2 (Invitrogen,Carlsbad,CA,United States) was used for the transfection of the cells,which was achieved by using LipofectamineTM RNAiMAX (Invitrogen,Carlsbad,CA,United States),according to the manufacturer's lipofection protocol.The sequence of siRNA for PKM2 was 5'-GCAGCUUUG AUAGUUCUGATT-3'.The nonspecific miRNA (Invitrogen,Carlsbad,CA,United States) was used as a control for nonspecific effects.The effects manifested after the introduction of the mature siRNA into the cells were assayed at 72 h after the transfection.
Statistical analysis
Data are expressed as the mean ± SEM.Statistical significance was determined by oneway ANOVA using SPSS 17.0 (SPSS,Chicago,IL,United States).P< 0.05 was considered statistically significant.
RESULTS
p-STAT3 participates in the formation of liver precancerous lesions in rats by activating hepatic progenitor cells
By HE staining,many heterocellular clusters in the liver parenchyma and portal area were observed in Solt-Farber model rats,the cells in the heterocellular clusters were characterized by deep nuclear staining,nuclear atypia,large karyoplasmic ratio,disordered arrangement of hepatocyte cords,and inflammatory cell infiltration.Compared with the model group,heterocellular clusters were hardly observed in the DEN + PH or normal group.In addition,significant proliferation of hepatic oval cells was observed in the liver tissues of rats both in the model group and AAF + PH group(Figure 1A).By immunohistochemistry,GST-π,a neoplastic marker in the early stages of carcinogenesis in HCC[28],was clearly stained in the heterocellular clusters in the model group (Figure 1B).In contrast,GST-π was hardly expressed in the DEN + PH,AAF + PH,or normal group,except in the portal area (Figure 1B).This result was also shown by Western blot (Figure 1C).Oval cell 6 (OV6) and cytokeratin 19 (CK19) are molecular markers of hepatic progenitor cells in rats.By immunofluorescence,OV6 was strongly expressed in the heterocellular clusters in the model group (Figure 1D),consistent with the studies of Solt and Dunsford[23,29].Western blot analysis also showed that the expression of CK19 in the model group was elevated (Figure 1F)[30].These results suggest that the hepatic progenitor cells are activated in liver precancerous lesions in rats.Then,we further evaluated p-STAT3 expression in liver precancerous lesions.The expression of p-STAT3 in the model group was significantly higher than that in the DEN + PH,AAF + PH,and normal groups (Figure 1D and E).Notably,OV6 and p-STAT3 were co-expressed in the same heterocellular clusters(Figure 1D).These results suggest that the activation of STAT3 is closely related to the activation of hepatic progenitor cells in the progression of liver precancerous lesions in rats.
The Warburg effect is increased in liver precancerous lesions in rats,and PKM2 in activated hepatic progenitor cells is associated with liver precancerous lesions
We detected glucose and lactate concentrations in liver tissue to determine whether the Warburg effect was initiated at the stage of liver precancerous lesions.The residual concentration of glucose in the model group was markedly decreased (Figure 2A).Likewise,the concentration of lactate in the model group was elevated (Figure 2B).These results illustrate that the Warburg effect has been initiated at the stage of the liver precancerous lesions in rats.
PKM2 plays a vital role in the Warburg effect of tumors[31].We detected the expression of PKM2 in liver precancerous lesions.The result showed that the expression of PKM2 in the model group was significantly higher than that in the DEN+ PH,AAF + PH,or normal group,and there were a large number of PKM2-positive cells around the portal area in the model group (Figure 2C and D).Especially,PKM2 was elevated in activated hepatic progenitor cells in the model group (Figure 2E).These results suggest that PKM2 is involved in the progression of activated hepatic progenitor cells in the liver precancerous lesions in rats.
p-STAT3 promotes malignant transformation of rat hepatic progenitor cells by enhancing cell proliferation
In order to explore the mechanism of STAT3 involvement in liver precancerous lesions,we replicated a model of transformed hepatic progenitor cellsin vitrousing MNNG and H2O2in light of previous findings[25,26].Transformed WB-F344 cells became anomalous and changed in size,and contact inhibition disappeared compared with wild WB-F344 cells.The rate of aneuploidy in transformed WB-F344 cells increased to 24% compared with wild-type WB-F344 cells (Figure 3A).Furthermore,the results of soft agar assay showed that clonal formation rate of transformed WBF344 cells was increased compared with the control group (Figure 3B).This indicates that the transformed cells had the characteristics of anchorage-independent growth,which is considered the most accurate and stringentin vitroassay for detecting malignant transformation of cells.AFP expression further supported the conclusion above (Figure 3C).These results suggest that the WB-F344 cells induced with MNNG and H2O2gained the characteristics of transformed cells.
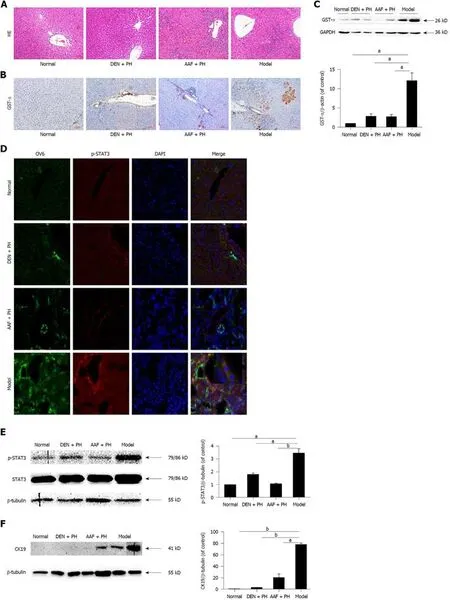
Figure1 Pathological changes in liver precancerous lesions in rats.
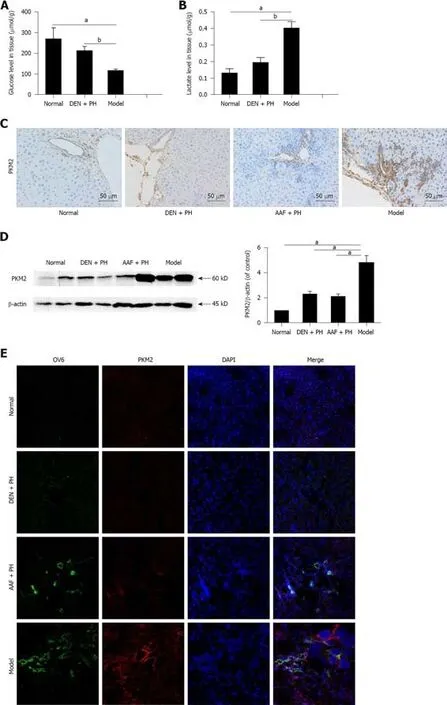
Figure2 The Warburg effect is initiated in liver precancerous lesions in rats.
In addition,p-STAT3 expression in transformed WB-F344 cells was higher than that in wild-type WB-F344 cells (Figure 3D).Next,we used stattic (MedChem Express,Monmouth Junction,NJ,United States),a specific inhibitor of STAT3,to suppress the activation of STAT3 in transformed hepatic progenitor cells.Compared with transformed WB-F344 cells,p-STAT3,aneuploidy,the clonal formation rate,and AFP expression decreased following 4 μmol/L stattic treatment (results of cell viability are not shown) (Figure 3A-D).These results indicate that STAT3 promotes malignant transformation of hepatic progenitor cells in rats.
To determine the mechanism by which STAT3 promotes malignant transformation of hepatic progenitor cells in rats,we measured PCNA expression and the cell cycle in transformed WB-F344 cells.As shown in Figure 3E,PCNA was highly expressed in transformed WB-F344 cells compared with wild-type WB-F344 cells,and PCNA was inhibited following stattic treatment.The percentage of transformed WB-F344 cells in S phase was increased compared with wild-type WB-F344 cells.STAT3 inhibition by stattic also significantly mitigated the increase in transformed WB-F344 cells in S phase (Figure 3F).These results suggest that STAT3 promotes malignant transformation of hepatic progenitor cells by enhancing cell proliferation.
STAT3 promotes the Warburg effect by enhancing p-PKM2 in transformed hepatic progenitor cells in rats
As shown in Figure 4A,transformed WB-F344 cells took up much higher amounts of glucose than wild-type WB-F344 cells.Similarly,lactate production in transformed WB-F344 cells was higher than that in wild-type WB-F344 cells (Figure 4B).These results suggest that the Warburg effect was initiated in transformed hepatic progenitor cells.The production of ATP in transformed WB-F344 cells was steady compared with wild-type WB-F344 cells (Figure 4C),in accordance with a previous study[32].In addition,in order to investigate the effect of PKM2 on the Warburg effect in transformed hepatic progenitor cells,we also measured the expression of p-PKM2.By Western blot analysis,it was discovered that the expression of p-PKM2 in transformed WB-F344 cells was significantly higher than that in wild-type WB-F344 cells (Figure 4D).The same results were obtained by immunofluorescence assay,and the expression of PKM2 in the nucleus was significantly increased in transformed WB-F344 cells (Figure 4E).
To determine the function and mechanism of STAT3 in the Warburg effect in transformed hepatic progenitor cells,we measured the influence of stattic treatment on the Warburg effect.The consumption of glucose,the production of lactate,and ATP levels were decreased following STAT3 inhibition by stattic (Figure 4A-C).These results indicate that STAT3 promotes the Warburg effect in transformed hepatic progenitor cells.And after STAT3 inhibition by stattic,p-PKM2 expression and nuclear translocation were also decreased (Figure 4D and E).These results indicate that STAT3 promotes the PKM2 expression and nuclear translocation in transformed hepatic progenitor cells.
To explore whether STAT3 works through PKM2,TEPP-46 (MedChem Express,Monmouth Junction,NJ,United States),a potent and selective activator of PKM2 that promotes the formation of PKM2 tetramers,was used to act on the malignant transformed WB-F344 cells after stattic treatment[33].The results showed that the suppressed Warburg effect by stattic was further reduced in transformed WB-F344 cells.Further use of PKM2 siRNA also resulted in the similar phenomenon (Figure 4A-C).These results suggest that STAT3 promotes the Warburg effect in transformed hepatic progenitor cells probably by upregulating p-PKM2.
DISCSSION
Recent studies have shown that STAT3 plays a vital role in the Warburg effect and adjusts the activity of key enzymes in the glycolytic pathway in tumor cells[34].STAT3 regulates aerobic glycolysis in breast cancer cells by stimulating hexokinase 2 transcription[14].STAT3 also increased glycolysis in myeloma cells by upregulating lactate dehydrogenase A and pyruvate dehydrogenase kinase 1[15].However,little work has been performed to determine the mechanism of STAT3 in promoting the Warburg effect in hepatocarcinogenesis.In our study,we observed that the Warburg effect was initiated at the stage of liver precancerous lesions,likely through activation of STAT3 and PKM2,in the Solt-Farber model.What’s more,STAT3 promoted the Warburg effect possibly by enhancing p-PKM2 in transformed hepatic progenitor cells in rats.
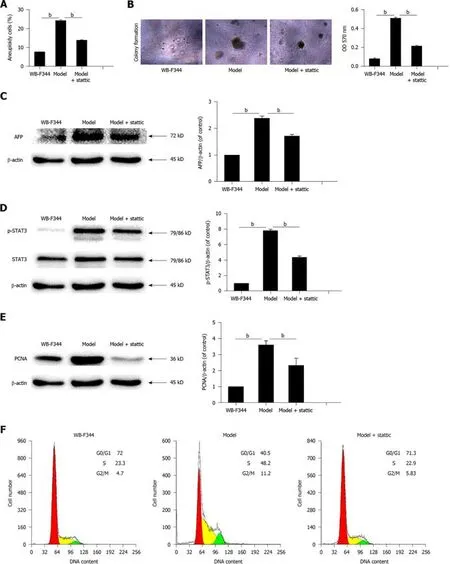
Figure3 Activated signal transducer and activator of transcription 3 promotes malignant transformation and cell proliferation in WB-F344 cells.
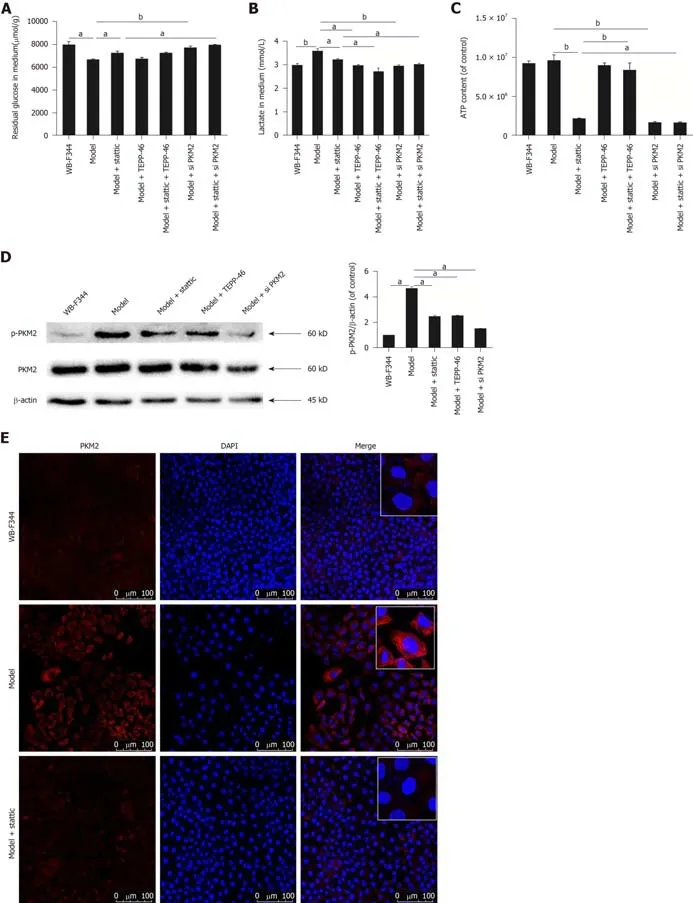
Figure4 Activated signal transducer and activator of transcription 3 promotes the Warburg effect by enhancing p-pyruvate kinase M2 in transformed WBF344 cells.
The theory of dedifferentiation of mature hepatocytes and the theory of abnormal differentiation of liver stem cells are two widely accepted hypotheses about the origin of liver cancer.The carcinoma might originate from mature liver cells or progenitor cells.Molecular studies have identified that adult hepatocytes are the source of HCC because they can directly degenerate into HCC following sequential genomic alterations.Alternatively,hepatic progenitor cells might generate HCC and intrahepatic cholangiocarcinoma with markers of progenitor cells[35],because hepatocytes and cholangiocytes each arise from a common progenitor (hepatoblasts)during liver development.Studies have noted a progenitor cell phenotype in many HCCs[36].And liver cancers with stem cell features have more aggressive clinical behavior and a worse prognosis than those without stem cell features.The Solt-Farber model is an established hepatic precancerous animal model.GST-π detoxifies electrophiles through conjugation to thiol-reduced glutathione (GSH) and is overexpressed in the early stages of carcinogenesis,including hepatic precancerous lesions.Our results confirmed that many heterocellular clusters were present in the liver parenchyma and portal area and that GST-π was highly expressed in liver precancerous lesions in Solt-Farber model rats (Figure 1C).In addition,our results revealed that OV6,a marker of hepatic progenitor cells in rats,was strongly expressed in the heterocellular clusters of the liver in the model group.As another marker of liver progenitor cells,the expression of CK19 was also elevated in the model group,suggesting that hepatic progenitor cells were activated (Figure 1D and F).
STAT3 is activated in HCC,but its effects in hepatic progenitor cells are not entirely clear.Our results revealed that p-STAT3 was strongly expressed in activated hepatic progenitor cells in the model group (Figure 1D and E),suggesting that STAT3 activation is involved in the stimulation of hepatic progenitor cells in the progression of liver precancerous lesions.
In a previous study,PKM2 promoted the Warburg effect in HCC,which provides the tumor with many advantages for cell growth and survival[37].However,the mechanism of PKM2 in the Warburg effect in HCC and liver precancerous lesions remains poorly understood.Our results indicate that the Warburg effect is initiated at the stage of liver precancerous lesions in rats (Figure 2A and B) and enhanced probably by PKM2 in activated hepatic progenitor cells (Figure 2C-E).
To further explore the mechanism of STAT3 in liver precancerous lesions in rats,we observed the effect of STAT3 in transformed WB-F344 cells induced with MNNG+ H2O2.Increased aneuploidy and anchorage-independent growth are vital feature of tumor cells[38].The size and shape of transformed WB-F344 cells changed,contact inhibition vanished,and aneuploidy,clonal formation rate,and AFP expression increased (Figure 3A-C),indicating that hepatic progenitor cells were transformed extensively.The transformed WB-F344 cells were treated with stattic,which inhibited the activation of STAT3,to observe the function of STAT3 during the malignant transformation of hepatic progenitor cells.The results showed that clonal formation rate,AFP expression,and aneuploidy fell significantly (Figure 3A-C),suggesting that STAT3 promotes malignant transformation of hepatic progenitor cells in rats.Intracellular STAT3 activation is closely associated with the cell cycle and proliferation in HCC cells.Our results also showed that STAT3 enhances malignant transformation of hepatic progenitor cells by promoting the expression of PCNA and accelerating G1/S phase transition (Figure 3E and F).
Dimeric PKM2 promotes cell proliferation and the Warburg effect in HCC[22],but its effects in hepatic progenitor cells are still unknown.Our results show that the Warburg effect was initiated and the p-PKM2 expression and nuclear translocation were increased in transformed WB-F344 cells (Figure 4A-E).This result implies that p-PKM2 probably promotes the Warburg effect in transformed WB-F344 cells.In addition,in the Solt-Farber model,the total protein expression of PKM2 in the liver was also higher,which may be due to the complexity of the internal environment in rats compared with that in cell culture.Following stattic treatment,p-PKM2 expression and nuclear translocation decreased,and the Warburg effect was suppressed (Figure 4A-E).Further use of PKM2 siRNA on the basis of stattic treatment in transformed WB-F344 cells further reduced the Warburg effect (Figure 4A-C).An interesting result is that,treatment with TEPP-46,a potent and selective activator of PKM2,in transformed WB-F344 cells after treatment with stattic,further reduced the production of lactate,increased ATP level,but had no significant impact on glucose consumption,which is consistent with previous studies[39,40].The reasons are that TEPP-46 promotes the formation of PKM2 tetramer and forms an enzyme with high pyruvate kinase activity as well as mediates the last step of glycolysis by promoting glycolysis flux,and then inhibits the formation of PKM2 dimer.That is to say,TEPP-46 in fact inhibits the Warburg effect.These results indicate that STAT3 promotes the Warburg effect possibly by enhancing p-PKM2 expression in transformed hepatic progenitor cells in rats.Previous studies have demonstrated that PKM2 promoted oncogenesis probably by phosphorylating STAT3[41].Hence,whether there is positive feedback between STAT3 and PKM2 in malignant transformation of hepatic progenitor cells remains to be further studied.
In summary,our results provide the evidence that the Warburg effect has been initiated at the stage of the liver precancerous lesions in rats,perhaps through high expression of PKM2 and p-STAT3 in activated hepatic progenitor cells in rats.Furthermore,STAT3 promotes the Warburg effect possibly by upregulating p-PKM2 expression and leading to malignant transformation of hepatic progenitor cells in rats.These results can help better understand the role of PKM2 and STAT3 in the formation of HCC.
ARTICLE HIGHLIGHTS
Research background
Hepatocellular carcinoma (HCC) is the fifth common malignant tumor worldwide and has a poor prognosis.HCC is closely associated with the potential precancerous lesions.Early treatment at the precancerous stage could significantly prevent the occurrence of HCC.Signal transducer and activator of transcription 3 (STAT3) and pyruvate kinase M2 (PKM2) can be activated and enhance the Warburg effect in HCC.However,whether activation of STAT3 enhances the Warburg effect in liver precancerous lesions in rats,and the relationship between STAT3 and PKM2 remain unclear.Hence,clarifying the mechanism of liver precancerous lesion of HCC are very important.
Research motivation
Investigating the mechanism of STAT3 and the Warburg effect in liver precancerous lesions in rats may suggest potential molecular mechanisms of hepatocellular carcinogenesis,and further offer the potential for developing novel therapeutic strategies for HCC treatment.
Research objectives
To measure the expression of STAT3 and PKM2,and the Warburg effect in liver precancerous lesions in rats and in transformed WB-F344 cells,and to investigate the possible molecular mechanisms of STAT3 in liver precancerous lesions.
Research methods
The Solt-Farber model is an established hepatic precancerous animal model.The transformed WB-F344 cells were induced with N-methyl-N’-nitro-N-nitrosoguanidine (MNNG) and hydrogen peroxide (H2O2).The contents of glucose and lactate in the tissue and culture medium of the cells were detected with a spectrophotometer.The protein levels were examined by Western blot and immunofluorescence.
Research results
Here,we provide the evidence that the Warburg effect was enhanced in liver precancerous lesions in rats,perhaps through high expression of PKM2 and p-STAT3 in activated oval cells.STAT3 activation promotes the Warburg effect by activating PKM2 in transformed WB-F344 cells induced with MNNG and H2O2.
Research conclusions
STAT3 activation promotes the Warburg effect by enhancing the phosphorylation of PKM2 in transformed WB-F344 cells.The Warburg effect,PKM2 and STAT3 expression were increased in liver precancerous lesions in rats.
Research perspectives
We have carried out some rat experiments andin vitrocell experiments,but further studies are needed to explore the mechanism of liver precancerous lesions.We also need to collect clinical samples for further validation.STAT3 and PKM2 may be potential diagnostic or therapeutic targets and used for clinical diagnosis and therapy in the future.
 World Journal of Gastroenterology2019年16期
World Journal of Gastroenterology2019年16期
- World Journal of Gastroenterology的其它文章
- Intra-abdominal desmoid tumors mimicking gastrointestinal stromal tumors — 8 cases: A case report
- Assessment of chronic radiation proctopathy and radiofrequency ablation treatment follow-up with optical coherence tomography angiography: A pilot study
- Dual energy computed tomography for detection of metastatic lymph nodes in patients with hepatocellular carcinoma
- Comparison of decompression tubes with metallic stents for the management of right-sided malignant colonic obstruction
- Role and mechanism of circ-PRKCI in hepatocellular carcinoma
- Immune response pattern varies with the natural history of chronic hepatitis B