
Role of calpain system in meat tenderness:A review
2018-05-22 07:18:10BhtJmesMortonSusnMsonAlElDinBekhit
食品科學(xué)與人類健康(英文) 2018年3期
Z.F.Bht,Jmes D.Morton,*,Susn L.Mson,Al El-Din A.Bekhit
a Department of Wine Food and Molecular Biosciences,Faculty of Agriculture and Life Sciences,Lincoln University,Lincoln,7647,Christchurch,New Zealand
b Department of Food Sciences,University of Otago,P.O.Box 56,Dunedin 9054,New Zealand
ABSTRACT Aging is a popular method used by meat industry for improving the sensory attributes of meat.Despite the advent of many novel technologies,aging has not lost its charm and is still widely used commercially as a post-mortem intervention for tenderization.Aging improves the tenderness of meat through disruption of the muscle structure by intracellular proteolytic systems.Muscles undergo various molecular changes that cause proteolysis of key myofibrillar and cytoskeletal proteins,disrupting the overall integrity of muscle cells.Although several endogenous proteolytic systems are capable of post-mortem proteolysis,a great body of scientific evidence supports a major role for the calpain system.Calpains are intracellular calcium-dependent cysteine proteases found in most eukaryotes.At least three calpains(μand m-calpains and calpain 3)and calpastatin,their specific endogenous inhibitor,are found in muscle.They are known to be involved in the proteolysis of functionally relevant structural proteins such as the myofibrillar proteins and cytoskeletal anchorage complexes.These ubiquitous proteases are also present in mitochondria and play important roles in a variety of pathophysiological conditions including apoptotic and necrotic cell death phenomena.This review discusses the role and contribution of the calpain system and the factors that influence calpain activity during aging.
Keywords:Aging Tenderness Calpains Proteolysis Calpain activity Factors
1.Introduction
Tenderness is one of the most important characteristics of meat and inconsistent tenderness is a particular issue in beef.After slaughter,the muscle undergoes various biophysico-chemical changes and events that converts it into meat.This process can be divided into three phases[I]the pre-rigor phase during which collagen content mainly contributes to the toughness,[II]rigor phase during which further toughening occurs due to muscle shortening,and[III]tenderization phase or resolution of rigor during aging during which the muscles undergo a series of changes and observe a remarkable improvement in tenderness.Thus,there are three major factors that are involved in the process that determine the tenderness of the meati.e.collagen content and solubility (background toughness),the extent of muscle shortening,and lastly the aging[1]and these vary with time,temperature,individual genotype,muscle types,and animal species[2].However,the common focal point to maximize tenderness is the appropriate activation of the endogenous proteolytic enzymes(calpains,cathepsins,caspases)responsible for degradation of muscle fibers.Several studies have provided convincing evidence that most of the tenderness development during aging is the result of post-mortem proteolysis of key structural proteins,including desmin,titin,nebulin,troponin-T and others [3,4].Microscopic studies have shown disruption of sarcomere structure with Z-disks degraded within a few days of aging[3].These structural changes caused by post-mortem proteolysis of myofibrillar proteins are the main cause of tenderization during aging.
2.Factors affecting the tenderness during aging
Post-mortem aging results in optimum improvements in the tenderness of meat,however,it does not ensure uniformity in the tenderness [5]as it is influenced by several genetic and environmental factors.
2.1.Temperature
The temperature of storage during aging is a crucial factor and several studies have shown a significant effect of rigor temperature on meat quality attributes[6,7].If it is below freezing point(-2 to-3°C),the activity of enzymes involved in the aging process will cease and if the temperature is higher,the enzymes will work effectively but there is a risk of microbial spoilage and other safety issues(i.e.pathogen growth).Further,the rate of glycolysis also increases in muscles with an increase in rigor temperature [8]which leads to a rapid decline in pH while the temperature of the muscle is relatively high causing conditions for protein denaturation.These conditions result in pale,soft,and exudative(PSE)-like meat characteristics [9,10].So,finding an appropriate aging temperature is very important and most of the studies report a temperature of around 0–4°C for aging.
The early post-mortem temperature of the muscle drastically affects the rigor shortening with a minimum degree of shortening in the range of 14–20°C.The maximum degree of shortening is observed when the muscle goes into rigor in the range of 0–10°C and this can result in a 50 percent reduction in the normal length of sarcomeres [11].It is well established that if the internal temperature of carcasses drops below 10°C and the pH is high(pH >6.2),it will result in toughening of the meat due to cold shortening[12,13].High rigor temperatures(>25°C)have also been reported to have an adverse impact on meat tenderness due to heat shortening[8,11,14,15].As much as 30 percent shortening has been reported when pre-rigor muscle was kept in the range of 20–40°C.The lowest amount of rigor shortening,around 10 percent,was observed when the pre-rigor muscle was kept in the range of 15–20°C[11].The temperature of the muscle during the pre-and post-rigor phase profoundly affects its metabolism [16]and the rate and extent of shortening observed at various temperature extremes is most likely determined by the effect of this temperature on muscle metabolism.These findings emphasize the importance of the chilling temperatures maintained during aging which can have serious implications on the tenderness of the meat.
Another factor that may aggravate the effect of temperature on tenderness and needs to be considered while designing chilling protocols is the variation in the rate of chilling due to differences in the size and composition of carcasses.Very large carcasses and carcasses with thick layers of fat experience a pronounced gradient of temperature with the innermost muscles experiencing the lowest cooling rate.One solution to counter this is stepwise chilling which allows rigor to proceed at comparatively higher temperature by initially maintaining a temperature of 10–15°C for up to 6 h and 4°C or less thereafter [17,18].This cooling system has been suggested to prevent cold shortening as well as limiting the microbial growth[17].
2.2.Time of aging
The duration of aging has a great bearing on the quality of the meat and varies considerably with the temperature of aging.An extended aging time has been reported to introduce favourable texture and flavour to the final product[19],however,the optimal time for dry aging for beef is 14 to 21 days and 7 to 10 days for wet aging at 0 to 1°C[20].Although,Smith[21]found a significant 17%reduction in shear force from day 14 to 35 of dry aging,showing that improvements were still occurring at least from an objective assessment standpoint,Campbell et al.[22]found no real advantages for extended aging past 14 days.Type of muscles and the rate with which the process occurs also affect the time of aging [23].Soft muscles,like the ones in loin and tenderloin,require shorter aging time than round muscles to become tender due to their lesser amount of connective tissues [24,25].In dry aging,the air velocity also affects the rate of aging,as a sufficient airflow is required to remove moisture from the meat.Higher air velocity leads to increased moisture loss causing more shrinkage and resulting in greater weight and trim losses[26].
2.3.pH
The rate and extent of pH decline is another early post-mortem factor that occurs with the development of rigor and can play an important role in the tenderization of muscle.While pH decline can affect the tenderness by influencing the shortening as it is partially associated with rigor development,particularly in the case of cold shortening,it can also affect the tenderness through other mechanisms [27,28].A high or low pH in beef at 3 h post-mortem has been reported to produce a less tender product than the beef that had an intermediate pH.A moderate rate of pH decline is thus considered beneficial for improved tenderness and has been reported in different studies involving electrical stimulation [29],different muscle fiber types [27]andin vitroevaluation [30].The pH influences the activity and stability of proteolytic enzymes of the muscle cells[17].
2.4.Fiber-type composition
Another factor that may affect the tenderization process during aging is the fiber type composition of muscles as fibers vary in their rate of use of ATP and their susceptibility to chilling temperatures.Muscles with a high number of oxidative fibers are more vulnerable to cold shortening due to the accelerated loss of calcium sequestering ability of the sarcoplasmic reticulum during rapid temperature decline.The rate of aging is reported to be faster in fast-twitch glycolytic muscles than in slow-twitch oxidative muscles[31]which could be explained by their higher ratio of calpain to its inhibitor,calpastatin,than in slow-twitch oxidative muscles[32].Slow-twitch oxidative muscles have the highest amounts of calpain present; however,they also have the highest amounts of calpastatin,which prevents proteolysis[33].The comparatively more developed sarcoplasmic reticulum and T-tubule system and thinner Z-bands of the fast-twitch glycolytic muscles are the other contributing reasons.The proteins in the Z-bands of the fast-twitch muscles are also reported to be more susceptible to early postmortem proteolytic degradation [34].The increase of fast-twitch glycolytic fibers has beneficial effects on the tenderness of beef during aging[35].An increase in the rate and extent of post-mortem pH decline with increased proportion of fast-twitch glycolytic fibers(type II fibers) in muscles has been reported by several studies,while a high proportion of type I fibers(oxidative fibers)decrease the rate and extent of pH decline[36],increase shortening of muscles,and influence the tenderness [37].Increasing the proportion of type IIB fibers in bovine muscles increased their tenderness due to improved post mortem aging whereas increasing the proportion of these fibers in pork muscles decreased both tenderness and water holding capacity.Longissimus dorsi,SemitendinosusandSemimembranosusamong bovine muscles have a high proportion of type IIB fibers [38,39],whereas a high degree of type I fibers is found inPsoas major,Biceps brachii,TrapeziusandBrachialis[38,40].
2.5.Sarcomere length
The influence of sarcomere length on the tenderness of meat has been studied extensively [41,42],however,there are separate views about its role on tenderness of the meat.While Rhee et al.[41]observed that overall sarcomere length was significantly correlated to tenderness;others[4,43,44]believed that the extent of proteolysis was independent of sarcomere length as aging was equally effective in improving the tenderness of shortened as well as non-shortened meat.However,several workers[45,46]have reported a reduction in the degradation of proteins,and subsequent improvement in tenderness,due to shortened sarcomeres because of reduced access to susceptible proteins for proteolysis.Many workers[44,45,47,48]have concluded that both post-mortem proteolysis and sarcomere shortening have a direct influence on meat tenderness,however,their interactions are not well understood.
2.6.Proteolysis
Tenderness of meat largely depends on the overall integrity of muscle cells that can be disrupted by the degradation of key myofibrillar and cytoskeletal proteins[49].Weakening of the myofibrillar structures by proteolytic enzymes due to the degradation of muscle proteins like titin,nebulin and desmin has been reported during aging by several workers [50,51].The meat tenderization mechanisms during aging are primarily recognized as enzymatic in nature and involve intracellular proteolytic systems capable of post-mortem proteolysis[52].Several endogenous proteolytic systems present in meat including calpains,the lysosomal proteases,and cathepsins have the capability of degrading myofibrillar and cytoskeletal proteins [47],however,the calpain system has been shown to be responsible for the proteolysis of cytoskeletal proteins(titin and nebulin) and intermediate filaments (desmin) during aging [50].There is a great body of research that links the calpain system to the tenderization of muscles and considers it responsible for the majority of post-mortem proteolysis and a significant driver of meat tenderization during aging[50,53–56].
Calpain system comprises endogenous proteases (calpains)which are considered as the primary candidates for muscle protein degradation initiated during the first 24 h post-mortem and their inhibitor,calpastatin [50,55,56].Considerable evidence supports the role of the calpain enzymes in proteolysis and tenderization in lamb,beef and pork.Studies suggest that different tenderization rates in different species are inversely correlated to the calpastatin:calpain ratio [57].These enzymes have been reported to cleave the same muscle proteins that are degraded during post-mortem aging[50,53,58]and have the capability to degrade specific muscle proteins including intermediate filament proteins,like desmin and the structural proteins,like titin [50,55]with minimal effects on myosin and actin [59].Post-mortem proteolysis,as measured by the loss of intact desmin and troponin-T,was limited in transgenic mice over-expressing calpastatin[60].These results were later confirmed by a similar study in μ-calpain knockout mice [53].Other evidence that supports the importance of calpains comes from analyzing the effects of β-adrenergic agonists which are involved in muscle protein turnover and have been reported to reduce the muscle protein degradation [61]by elevating the calpastatin activity resulting in tougher meat[62].Likewise,the high levels of calpastatin found in the hypertrophied muscles of callipyge lambs was associated with a significant reduction in post-mortem proteolysis and meat tenderness [63].A simple injection of postmortem muscle with calcium accelerates postmortem proteolysis and tenderization[64,65],suggesting the involvement of calpains.All this evidence indicates a role for the calpain system in post-mortem proteolysis and tenderization during aging,however,there are several different calpain isoforms which may be responsible for the protein degradation.
3.The calpain system
The calpain system is the most extensively studied enzyme system involved in meat tenderization[54,66].They are a large family of cysteine proteases which are present in almost all eukaryotes and a few bacteria.They are unusual proteases in that they require calcium for their activity.They are intracellular proteinases with optimum activity at neutral pH.The system comprises several isoforms of the proteolytic enzyme calpain and their endogenous inhibitor,calpastatin[67,68].The μ-calpain and m-calpain are the two best-characterized isoforms out of fifteen genes for calpain isoforms that have been identified in the human genome and these have been classified on the basis of tissue distribution into ubiquitous and tissue-specific forms[68,69].The calpain system in the skeletal muscles consists of two ubiquitously expressed proteasesviz.μ-calpain and m-calpain in addition to p94(tissue specific calpain 3)and calpastatin[54,68,70].The μ-calpain is mostly bound to myofibrils(70%)and most of the m-calpain is located in the cytosol[71,72]whereas the location of their inhibitor,calpastatin,coincides with them[73].The μ-calpain and m-calpain require different Ca2+levels for their activation.While m-calpain needs 400–800 μM Ca2+for half-maximal activity,μ-calpain needs only 3–50 μM Ca2+for half-maximal activity[68].
3.1.Structure of calpains
Fig.1 shows the domain structures of μ-calpain,m-calpain and calpastatin.These calpains have highly homologous structure and both μ-calpain and m-calpain are heterodimers composed of a similar large 80 kDa catalytic subunit and share an identical 28 kDa small subunit[74].The large 80 kDa subunit of each calpain is composed of four (I,II,III,IV) domains [68].The small subunit has two domains (V,VI).Domain V is rich in glycine and is the site for phospholipid binding.Domain VI contains five Ca2+-binding sites also known as EF-hand motifs[75].The N-terminal domain of 80 kDa subunit,domain I,has no sequence homology to any known polypeptide and its removal modulates the proteolytic activity.The catalytic domain,domain II,contains a cysteine residue (subdomain IIA)as well as a histidine residue(sub-domain IIB)that are in relative positions that are conserved in all cysteine proteinases[74].The activity of calpains may depend on other regions of the molecule in addition to the active site,as studies have confirmed that the fragments containing the catalytic domain(domain II)may not have catalytic activity by themselves[76]showing the importance of conformational state on the activity of these enzymes[77].A Ca2+-binding domain,domain III,is not homologous to any other known protein and is linked to the catalytic domain II.It has sequences that predict EF hand calcium binding sites[68]and may regulate the activity of calpain through binding of phospholipids and critical electrostatic interactions [77,78].A calmodulin-like domain,domain IV,is also known as penta-EF domain and has five EF-hand calcium binding sites.The first four EF-hands in domain VI also contain Ca2+-binding sites.The fifth EF-hand motif in the carboxyl end of IV and VI domains do not bind calcium and interact with each other to form a heterodimer of the two calpain subunits[11,79].
Calpastatin,an endogenous inhibitor of μ- and m-calpain,is a 70–80 kDa protein and has an N-terminal L domain and four repeating domains(I,II,III,IV).Each of the four repeating domains is able to inhibit one calpain molecule[80,81].Calpastatin in the erythrocytes is a small 46 kDa version of the protein and some isoforms,particularly in cardiac muscle,also contain a 68-amino acid XL Nterminal domain[81,82].
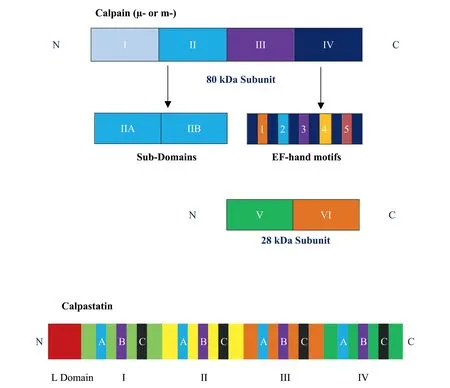
Fig.1.Domain structures of μ-calpain(calpain 1),m-calpain(calpain 2)and calpastatin(Not to scale).
Originally referred to as p94,calpain 3 is a 94 kDa calpain isoform bearing sequence homology of approximately 50% with the large domain of μ-and m-calpain[83].It may form homodimersin vivoas it lacks the 28 kDa small subunit [84].This calpain protease is specific to skeletal muscle and is also found in an 82 kDa form in the retina and lens[69].
3.2.Mechanisms of action
Both the calpainsi.e.μ-calpain and m-calpain bind calcium ions at various locations during activation.Although,m-calpain can bind 11–20 calcium ions and μ- calpain may bind 5–8 calcium ions,a structural study of m-calpain in the presence of 5 mM calcium has shown that it only binds 10 calcium ions [85].Depending on the calcium availability,the EF-hand motifs of IV and VI domains can bind three to four calcium ions which cause a small conformational shift that results in the dissociation of small subunit [86].However,this conformational change caused due to binding of calcium ions to the EF hands is probably not enough to result in activation of m-calpain[87].The protease core domains,IIA and IIB,have no protease activity by themselves and it is only the presence of calcium ions on each one binding site that causes a conformational shift which allows protease activity [88].Thus,proteolysis is dependent on the binding of calcium ions at the two protease core sites which disrupts the salt bridges between the Glu333 and Arg104 residues in domain II,allowing the catalytic triad(cysteine(Cys105)residue in domain IIA and asparagine(Asp262)and histidine(His286)residues in domain IIB)to come together,resulting in activation[69,89].Several factors,like phospholipid binding,autolysis,or binding to membrane activator proteins,have been reported to reduce the calcium requirement[90,91].
Unlike other endogenous proteases,like cathepsins,μ and mcalpain are quite specific for their substrates.They often cut their substrates at only a few sites and do not completely degrade the substrates to small peptides.The evidence is that their specificity is mostly determined by protein conformation rather than amino acid sequence[92,93].However,studies involving computer modelling have identified many substrate sequence specificities[94].
The inhibition of the activity of μ-calpain and m-calpain by calpastatin is also a calcium dependent event as calpain-calpastatin binding requires calcium.Calpastatin inhibits calpain by preventing the calpain proteolytic activation,membrane binding and the expression of catalytic activity [11].Conformational changes are induced in the calpains due to the binding of calcium ions,specifically in domains I-IV and VI.The region A and region C of the calpastatin molecule binds to domain IV and VI of calpain molecule,respectively.This causes the region B of the calpastatin inhibitory domain to make various contacts with regions of domains I to III of calpains,thus blocking the active site of calpains and inhibiting its activity [11,95].TIPPXYR is the central inhibitory sequence in the calpastatin molecule [95]and most of the isoforms of calpastatin vary in the XL or L domain,and thus do not show much change in the calpain inhibitory ability[68,96].
3.3.Properties of calpain isoforms and their role in proteolysis
3.3.1.μ-calpain and m-calpain
Several studies provide compelling evidence that tenderization of meat during post-mortem aging is predominantly modulated by the calpain system [51,53,54,97–99].Although,both isoforms(μ-calpain and m-calpain)target and cleave the same myofibrillar proteins,the calcium requirements for full activation of m-calpain is far higher than the calcium concentrations attained post-mortem(maximal 210–230 μmol/L)and will only be obtained if exogenous calcium is added to the meat[53,54].Therefore,μ-calpain is considered to be of much greater significance for tenderness development through postmortem proteolysis[98,100].Further,the activation of μ-calpain coincides with the period of post-mortem(within 3 days of slaughter) when proteolysis of key myofibrillar proteins takes place whereas m-calpain is not activated early post-mortem[101].The autolysis of μ-calpain and loss of most of the activity has been reported to occur within 7 days post-mortem [102]whereas mcalpain activity did not decrease in lambBiceps femorisup to 56 days post-mortem[63].Moreover,the degradation patterns similar to those observed in post-mortem muscle are produced when myofibrils are directly incubated with μ-calpain [50].Very little post-mortem proteolysis is observed in the muscles of μ-calpain knockout mice[53],suggesting its predominant effect on catalysis of proteins during postmortem aging[103].
Calcium is required for calpain activation,however,both μcalpain and m-calpain will also autolyze when exposed to the calcium levels necessary for their activation,thus proteolytic ability is accompanied with autolysis [11].The autolysis results in progressive degradation of 80 kDa subunit of μ-calpain to 78 kDa and eventually to 76 kDa whereas the mass of the 80 kDa subunit of m-calpain reduces to 78 kDa [104].The 28 kDa subunit of both μ-calpain and m-calpain is reduced to 18 kDa after losing its glycine-rich domain [105].The calcium requirement of μ-calpain and m-calpain for their activity is reduced by brief autolysis whereas extended autolysis results in inactivation of these enzymes[106].This reduction of the calcium requirement explains how these enzymes are active in cellular conditions which rarely,if ever,acquire the calcium levels needed for their activity.The autolyzed forms of both μ-calpain(76 kDa)and m-calpain(78 kDa)requires 92 percent and 82 percent less calcium,respectively,than their unautolyzed (80 kDa) forms [106]although,both the forms of the enzymes have been shown to have activity.The autolyzed form of μ-calpain binds tightly to myofibrils and other subcellular organelles due to its increased hydrophobic nature[107].Since μand m-calpain activity and proteolysis is always accompanied by autolysis,the presence of autolyzed form of calpain in post-mortem tissue means it has been active and measurement of calpain autolysis gives an estimate of the activity of these enzymes up to the analyzed point of time.
3.3.2.Calpain 3
Discovered as the third calpain and named so,calpain 3 is not inhibited by calpastatin [108]and requires only nanomolar levels of calcium for activation [109].In addition to protease activity,it also seems to play a structural role in skeletal muscle fibers in the triad regions composed of one T-tubule and two terminal cisternae[110].This structure regulates calcium release during contraction and lack of calpain 3 was reported to decrease the calcium release in the fibers due to an impairment of the triad-associated protein complex [110].Expressed almost exclusively in skeletal muscle[83],this calpain is associated with the giant myofibrillar protein titin,also known as connectin.Calpain 3 has also been reported to degrade calpastatinin vitroand may possibly regulate the activity of μ-calpain and m-calpain if the same were truein vivo[108].Calpain 3 binds with titin at N2 or M-line region[111,112]which is a known site linked with proteolysis during meat tenderization [113].The binding of calpain 3 with titin protects it from auto-degradation and its removal from titin results in rapid autolysis.Due to this property the early investigations involving its purification were difficult.Calpain 3 is also susceptible to sodium-dependent autolysis and has been reported to have sodium-dependent activity with different substrate specificity compared to its calcium-dependent activity[114].
Although calpain 3 was initially thought to have a major role in the post-mortem proteolysis and tenderization process due to its association with titin,studies over the years have nullified its importance and the role calpain 3 may play in the process,if any,appears to be minor.Given the fact that calpain 3 is not inhibited by calpastatin[108],animals with very high calpastatin would not produce tough meat had it have any substantial role in meat tenderization process.Further,no association was found between calpain 3 expression and the tenderness level in porcineLongissimus dorsi[115].However,variations in tenderness in sheep have been reported to correlate strongly with the variations in calpain 3 mRNA and protein levels[116].To see the role of calpain 3(p94)in meat tenderization,Geesink et al.[117]compared the post-mortem proteolysis patterns of p94 knockout mice with wild-type mice and reported no detectable difference in desmin,nebulin,troponin-T or vinculin degradation,suggesting the non-involvement of calpain 3 in meat tenderization.
3.3.3.Proteolysis of desmin and troponin-T
There is a particularly well-defined organization of intermediate filaments in the skeletal muscles,which build a network that interconnects the myofibrils to each other at the Z-disks.In mature muscle fibers,this intermediate filament network is mainly composed of the protein desmin [118]and provides mechanical strength to the muscle fibers by maintaining structural and functional integrity.These intermediate fibers extend and encircle myofibrils constituting a three-dimensional scaffold around the Z-disk and connect the entire contractile apparatus to the sarcolemma,the nuclei,mitochondria,and other organelles [119].Desmin proteolysis is one of the earliest markers for mammalian skeletal muscle during aging as it participates in the early establishment of sarcomere structure.Due to its structural functions,desmin degradation is believed to be essential for the tenderization of meat [3]and is believed to have a significant impact on meat quality during post-mortem aging[3,50].
Several workers have reported the degradation of desmin during meat aging[50,120–124].Degradation of desmin has been directly linked to calpain activity usingin vitromodels of post-mortem muscles [50,125–127].Both m- and μ-calpains have been reported to effectively degrade desmin at identical sites[67,126].Degradation of desmin during early post-mortem aging was reported to generate fragments from 50 to 39 kDa in pork[120,128].Degradation of desmin was evident at 3 days post-mortem aging and after 7 days,it was almost completely degraded[129,130]and thus it is believed to have a significant influence on meat tenderization by affecting the shear force and water-holding capacity of meat[3,11,122].
Troponin-T subunit is the tropomyosin-binding component which makes up the elongated portion of troponin complex which is involved in the calcium-dependent regulation of skeletal muscle contraction.Due to its close relationship with meat tenderness,degradation of troponin-T,especially the 30 kDa fragment,is considered to be an indicator of meat tenderization [131].There is a good correlation between the two events as the degradation of troponin-T progresses simultaneously with the tenderization of meat during post-mortem aging[132].
Degradation of troponin-T possibly leads to the fragmentation of the myofibrils by disrupting its interaction with other actin filament components.Like desmin,degradation of troponin T has also been directly linked to calpain activity usingin vitromodels[50,125–127]and is used in proteomics as a marker for the ongoing proteolysis in beef [133]and in pig meat [134].Polypeptide segments of 28 and 30 kDa are obtained from the degradation of intact troponin-T by μ-calpain.The magnitude of these concomitant polypeptides strongly correlated with the meat tenderness during postmortem aging[135].Thus,troponin T degradation has been used in pioneering studies aimed at meat tenderness prediction[136].
3.4.Factors affecting the activity of calpains
In addition to the sarcoplasmic Ca2+concentration,several environmental factors like temperature,pH,and oxidation have been reported to influence the activity of calpains[53,56,59].
3.4.1.Temperature
Influence of elevated temperature during early post-mortem on the proteolysis and calpain activity has been reported by several studies [7,137–139].The μ-calpain is very sensitive to temperature [140]and its activity has been observed to be primarily regulated by muscle temperature in post-mortem bovine muscle[141].Accelerated μ-calpain activation has been reported in lamb as well as beef muscles when incubated at elevated temperatures[7,139,142].Faster μ-calpain activation was observed by[7]at temperatures of 25–35°C in lamb and by[139]at 36°C in beef by using immunoblot analysis of muscle incubated for 24 h.Likewise faster proteolysis was observed in pieces of beef muscle incubated at 25°C for 8 h post-mortem [138].Although a faster proteolysis was also observed in beef muscles that entered rigor at 15°C up to 3 days post-mortem [137],less total proteolysis has been reported during prolonged aging [7,137]due to the lower stability of calpain at high temperatures causing early loss of μ-calpain activity[143].Elevated temperatures (above 25°C) during rigor have also been reported to decrease the activity of calpastatin[7].
The activity of m-calpain has been reported to be significantly faster at elevated temperatures[59,142]and this was attributed to both the increased temperature and the increased free calcium concentrations[69].The sarcoplasmic calcium level has been reported to increase in porcine muscle under certain time-temperature combinations to the level required for activation of μ-calpain and later also for m-calpain[144].An increase in extractable free calcium was also observed during the onset of rigor in beef muscles incubated at a temperature of 36°C[139].
3.4.2.pH
The pH is another factor that may affect the activity of calpains and proteolysis of their myofibrillar protein substrates.At pH values and ionic strengths physiologically found in postmortem muscle,both μ-calpain as well as m-calpain show less activity against myofibrillar protein substrates than at pH 7[5,55].Change in pH and ionic strengths may cause conformational changes in enzymes as well as in their substrates affecting the activity of enzymes and the susceptibility of substrates.These changes may render the substrates less vulnerable to cleavage by μ-calpain[55].The rate of decline of early postmortem pH is an important factor that influences the autolysis and activation of μ-calpain,with accelerated decline considered favourable for accelerated proteolysis of known calpain substrates [27,30].Slightly accelerated pH decline in the muscles may also lead to an accelerated rate of tenderization and,sometimes also have an advantage in water holding capacity[27,29].Too rapid pH decline is not considered favourable as it will reduce the autolysis of calpains and proteolysis of their substrates.In pork,a rapid pH-decline almost completely arrested autolysis of μ-calpain [145]and degradation of desmin and talin[146].Increased protein denaturation due to low pH and high temperature combinations may be responsible for this condition.
3.4.3.Oxidation
Oxidation can also affect the activity of μ-calpain and influence post-mortem tenderization.Incubation with H2O2(200 mmol/L)results in loss of activity of μ-calpain,reversible under reducing conditions,due to the formation of a disulfide bond between the active site Cys 115 and Cys 108 [147].It has also been reported to decrease the degradation of desminin vitro[30].Chen et al.[148]studied the effect of oxidation on the susceptibility of purified desmin to degradation by μ-calpain and reported that oxygen radicals changed the secondary structure and proteolytic susceptibility of desmin and decreased its degradation rate after incubation with μ-calpain for 1 h.While studying the effect of oxidation on the degradation of myofibrillar proteins by μ-calpain,Xue et al.[149]came up with the finding that oxidative modification of myofibrillar proteins changes their susceptibility.The oxygen radicals increased the proteolytic susceptibility of MHC and α-actinin and reduced the degradation of troponin-T.Irradiation,which induces protein oxidation,has also been reported to decrease μ-calpain autolysis and degradation of troponin-T and desmin in beefLongissimus[150].
4.Conclusions
Aging is an effective and traditional way of improving tenderness and other characteristics of meat.The tenderization effect of aging is primarily enzymatic in nature and involves activation of endogenous proteolytic enzymes which are responsible for degradation of muscle fibres.Several endogenous proteolytic systems available in the muscle are capable of proteolysis,however,the calpain system is generally identified as the main driver for most of the proteolysis and tenderization process.There are several factors,such as temperature,pH,and oxidation,that affect the activity of calpains and the tenderization process during aging.While aging is practiced extensively in meat industry because of its benefits and a superior final product,it also has some economic considerations in terms of time,space,labour and energy.
Conflict of interest
The authors declare that there is no conflict of interest.

- 食品科學(xué)與人類健康(英文)的其它文章
- Behavioral assessment of hippocampal function following dietary intervention
- Study of interaction between metal ions and quercetin
- Prophylactic effect of Kudingcha polyphenols on oxazolone induced colitis through its antioxidant capacities
- Nutraceutical support for respiratory diseases
- Corn phytochemicals and their health benefits
- GUIDE FOR AUTHORS