
The miR-183~96~182 cluster promotes tumorigenesis in a mouse model of medulloblastoma
2013-10-27 09:21:06ZengdiZhangSanenLiStevenChengDepartmentofDevelopmentalGeneticsNanjingMedicalUniversityNanjingJiangsu210029ChinaReceived25January2013Revised17February2013Accepted08June2013Epub10July2013
Zengdi Zhang, Sanen Li, Steven Y Cheng Department of Developmental Genetics, Nanjing Medical University, Nanjing, Jiangsu 210029, China.Received 25 January 2013, Revised 17 February 2013, Accepted 08 June 2013, Epub 10 July 2013
INTRODUCTION
Medulloblastoma is a tumor of the cerebellum and the most common malignant pediatric brain tumor.Medulloblastoma occurs most frequently in children between the ages of 5 and 10 years but may occur in adults as well[1-3]. Although 5-year in overall survival rates have reached 60%-80%, the prognosis for many medulloblastoma patients remains bleak. Many patients who develop the disease die from it and survivors, after conventional treatment regimens (surgery,radiotherapy, and chemotherapy), often face a variety of long-term neurological, neuroendocrine, and social sequelaes[2-4]. It is therefore imperative for us to gain a deeper understanding of the molecular pathogenesis driving medulloblastomas, in order to improve approaches for treating medulloblastomas.
MicroRNAs (miR)are small endogenous noncoding RNAs that play important roles in many biological processes, including tumorigenesis[5]. These 21-23 nucleotide (nt)RNAs are matured through a 2-step biogenesis mechanism from long, RNA polII transcripts[6]. Mature microRNAs are incorporated into the RNA-induced silencing complex, where they anneal to their recognition sequences in the 3‘-UTR of mRNA genes to attenuate gene expression through translational repression or mRNA degradation[7]. Several genomic studies have revealed the differential expression of microRNAs across various molecular subgroups of medulloblastoma. As previously reported,the miR-17/92 polycistron was identified as a putative medulloblastoma oncomir and associated with the sonic hedgehog-driven medulloblastoma compared with other subgroups[8,9].
In different comprehensive analyses of the human and mouse medulloblastoma oncomir, the miR-183~96~182 cluster is upregulated in several subgroups, including particularly aggressive clinical course involving high rates of metastasis and overall poor survival[10]. Based on the studies of cell lines, Weeraratne et al. found that the knockdown of the full miR-183~96~182 cluster results in the enrichment of genes associated with apoptosis and dysregulation of the PI3K/AKT/mTOR signaling axis. However, the specific role of miR-183~96~182 cluster in medulloblastoma and cerebellar development has not been described[11,12].
Cerebellar granule neuron progenitors (CGNPs)are proposed cells of origin for a subset of medulloblastomas. CGNPs undergo rapid Shh-dependent expansion perinatally in mice and humans, and excessive Shh pathway activity promotes medulloblastoma[13,14]. We used the Ptch1+/-; PtenFloxp/+; GFAP-Cre mouse model of medulloblastoma to study the effect of the PI3K/Akt signaling pathway activated on medulloblastoma tumorigenesis. We found that the miR-183~96~182 cluster was highly expressed in this Pten and Ptch1 loss associated medulloblastoma and promote tumorigenesis by controlling CGNP proliferation.
MATERIALS AND METHODS
Animal
Ptch1+/-mice were purchased from Jackson Laboratory[15]. C57BL/6 and PtenFloxp/Floxpmice were purchased from the Model Animal Research Center of Nanjing University[16]. GFAP-Cre mice were gifts from Minsheng Zhu's laboratory (Nanjing University, Nanjing, Jiangsu, China). PtenFloxp/Floxp mice were crossed with Ptch1+/-mice. The,n Ptch1+/-;PtenFloxp/Floxpoffspring mice were crossed with GFAPCre mice to obtain Ptch1+/-; PtenFloxp/+; GFAP-Cre mice. Mice were observed for symptoms of medulloblastoma at least twice-weekly for 12 months.Transgenic expression patterns of GFAP-Cre were examined with a R26R reporter line (129S4/SvJaeSor-Gt(ROSA)26Sortm1(FLP1)Dym, (Jackson Laboratory,Bar Harbor, Maine, USA)carrying a lacZ gene whose expression requires excision of loxP-flanked stop sequences. All mice were housed in a specific pathogen free animal room. The study protocol was approved by the local institutional review board at the authors'affiliated institutions. Animal welfare and the experimental procedures were carried out strictly in accordance with the Guide for Care and Use of Laboratory Animals (National Research Council of USA, 1996).
Histological study
For histological analysis, mice were perfused with PBS followed by 4% paraformaldehyde. The cerebella were removed, fixed in 4% paraformaldehyde overnight, and transferred to gradient ethanol for dehydration and embedded in paraffin. Sections (5 m)were stained with hematoxylin and eosin (Sigma, St Louis,MO, USA). Frozen sections (20 mm)were stained for β-galactosidase according to a previously reported method[17].
RT-PCR
Total RNA from mouse tissues was extracted using the RNAiso reagent (TaKaRa, Osaka, Japan).Stem-loop RT-PCR primers were designed as previously reported[18]. The U6 RNA was used for normalization. Stem-loop RT primers and PCR primers were synthesized by Invitrogen (Carlsbad, CA,USA). Reverse tran scription was performed using the PrimeScript? RT reagent Kit with gDNA Eraser(TaKaRa). The cycling condition of PCR was 95°C for 5 minutes, followed by 30 amplification cycles of 95°C for 15 seconds and 60°C for 1 minute. Stemloop RT primers sequences for these microRNA were listed as follows: mmu-mir-183-RT: 5'- GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGTGAA-3'; mmu-mir-96-RT:5'-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCAAA-3'; and mmumir-182-RT: 5'-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCGGTGT-3'.Forward PCR primers sequences for these microRNAs are: mmu-mir-183-FR: 5'-AGCCGTATGGCACTGGTAGAA-3'; mmu-mir-96-FR: 5'-AGCCGTTTGGCACTAGCACATT-3'; mmu-mir-182-FR:5'-AGCCGTTTGGCAATGGTAGAACTC-3'.
Culture of CGNPs
Granule neuron progenitors were purified from the cerebellum of 7-day-old C57BL/6 pups by using a modified protocol as previously reported[19,20]. Briefly,postnatal cerebella were triturated into single-cell suspensions that were loaded onto a step gradient of 35%and 65% Percoll (Amersham-Bioscience, Uppsala,Sweden)and separated by high-speed centrifugation for 10 minutes at 4°C. Granule cells and precursors were harvested from the 35/65% interface, washed in PBS/DNase, and further purified by depleting adherent cells with 2-hour incubation on normal tissue culture dishes. Their purity (> 95%)was assessed by immunostaining with markers of neurons and glia. Cells(5×105per well)were plated in four-well Lab-Tek chamber slides precoated with 100 μg/mL poly-D-lysine (Millipore, Billerica, MA, USA)and Matrigel(Beckton Dickinson, Bedford, MA, USA)in neural basal medium supplemented with 0.45% glucose,SPITE (Sigma), oleic acid albumin/linoleic acid (Sigma), B27 (Invitrogen), and N-acetyl cysteine (Sigma).
Retrovirus production and infection
To generate a mmu-miR-183~96~182 cluster expression clone, the micoRNA cluster was PCR-amplified from genomic DNA of C57BL/6 mouse and cloned into pENTR/D-TOPO plasmid (Invitrogen),then transferred into MSCV-P1G-gateway vector(Clontech, Mountain View, CA, USA)by Gateway LR Clonase II enzyme mix (Invitrogen). When CGNPs became attached, 250 μL medium was removed and 250 μL medium containing virus was added. Infection was repeated 3 times with 2 hours each time. The next day, the medium was changed for fresh complete medium with or without 10% ShhN conditional medium(ShhN CM), which was generated by transfecting 293T cells with a ShhN expression construct[21].
EdU staining
Identification of purified granule cells in the S phase was performed by incubation with Click-iT EdU Alexa Fluor 555 Imaging Kit (Invitrogen). EdU was added for 12 hours before fixation. EdU detection was performed according to the manufacturer's instructions (Invitrogen). Stained chambers covered with mounting medium containing DAPI (Sigma)were enumerated under a microscope. TIFF images of 4 random fields were taken for each experimental group by using the 20 × objective lens. The counting of EdU-positive and DAPI-positive CGNPs number was quantified by using ImageJ software. The kinetics of the proliferative response to mmu-miR-183~96~182 was analyzed by using GraphPad Prism 5.
Statistical analysis
CGNPs proliferation at each time point was measured as the mean ratio (EdU-labeling to DAPI-labeling cell number)from 4 random fields. Statistical significance of proliferation was determined by using Student's t-test.
RESULTS
Heterozygosity forPtenpromotes medulloblastoma in cooperation with Hedgehog signaling pathway
The founders of GFAP-Cre and their offspring were identified by PCR genotyping. Transgenic expression patterns of Cre were examined with a Rosa26 reporter line, 129S4/SvJaeSor-Gt(ROSA)26Sortm1(FLP1)Dym, carrying a lacZ gene whose expression requires excision of Floxp-flanked stop sequences.β-galactosidase staining revealed GFAP-Cre mediated recombination both in astrocytes and immature precursor cells of the external granular layer (EGL)in the developing cerebellum of 7-day old mice (Fig. 1A).To explore the role of Pten loss in the background of Ptch1+/-mice, we administered the GFAP-Cre transgenic mice to delete Pten in CNS lineages by using conditional knockout mice PtenFloxp/Floxp. Broad CNS deletion of Pten results in lethal hydrocephalus in early postnatal life; modeling efforts henceforth emphasized the PtenFloxp/+GFAP-Cre genotype.
In PtenFloxp/+mice, part of Ptch1 exon 1 (including putative start codon)and all of exon 2 were replaced with lacZ and a neomycin resistance gene in one allele[15]. Approximately 17.6% (3 out of 17 PtenFloxp/+)of mice expressing one allele of Ptch1 developed symptoms of medulloblastoma by 1 year of age. Most of these mice died 5 months after birth[22,23]. However, PtenFloxp/+PtenFloxp/+GFAP-Cre mice died of medulloblastoma from 2 months after birth. We found that, between 2 to 8 months of ages, 18/23 (78.2%)of the PtenFloxp/+PtenFloxp/+GFAP-Cre mice presented with acute-onset neurological symptoms. Similarly to Ptch1+/-mice, these mice began to display signs of illness, including domed head, hunched back, abnormal gait and/or decreased movement (data not shown).Histopathologically, these mice harbored malignant medulloblastoma. Tumor cells, regardless of whether the mouse was Ptch1+/-or Ptch1+/-PtenFloxp/+or GFAP-Cre, arose from the surface of the cerebellum and invaded the cortex and deep white matter of the cerebellum (Fig. 1B, panel a and b). The tumors in the Ptch1+/-mice showed small round or ellipsoid cells with a high nuclear-to-cytoplasmic ratio and round-to-oval or triangular hyperchromatic nuclei,corresponding to patterns of classic medulloblastomas in humans (Fig. 1B, panel c). In contrast, the tumors from Ptch1+/-PtenFloxp/+GFAP-Cre mice had a biphasic architecture that nodular areas consisted of differentiated cells with round nuclei and less condensed chromatin, and were intermingled with the undifferentiated hyperchromatic cells (Fig. 1B, panel d). The nodular areas were surrounded with desmoplasia, a pericellular deposition of collagen in this context, similarly to the appearance of desmoplastic medulloblastomas in humans. However, none of the PtenFloxp/+GFAP-Cre mice developed medulloblastoma. Therefore, in cooperation with the hedgehog signaling pathway, heterozygosity for Pten promoted medulloblastoma. Medul-loblastomas induced by mutations in the Shh pathway has been reported to originate from long-lasting proliferative CGNPs and are located within the cerebellar hemispheres[13,24,25]. Therefore, we speculated that the Ptch1+/-PtenFloxp/+GFAP-Cre medulloblastomas arise from CGNPs.
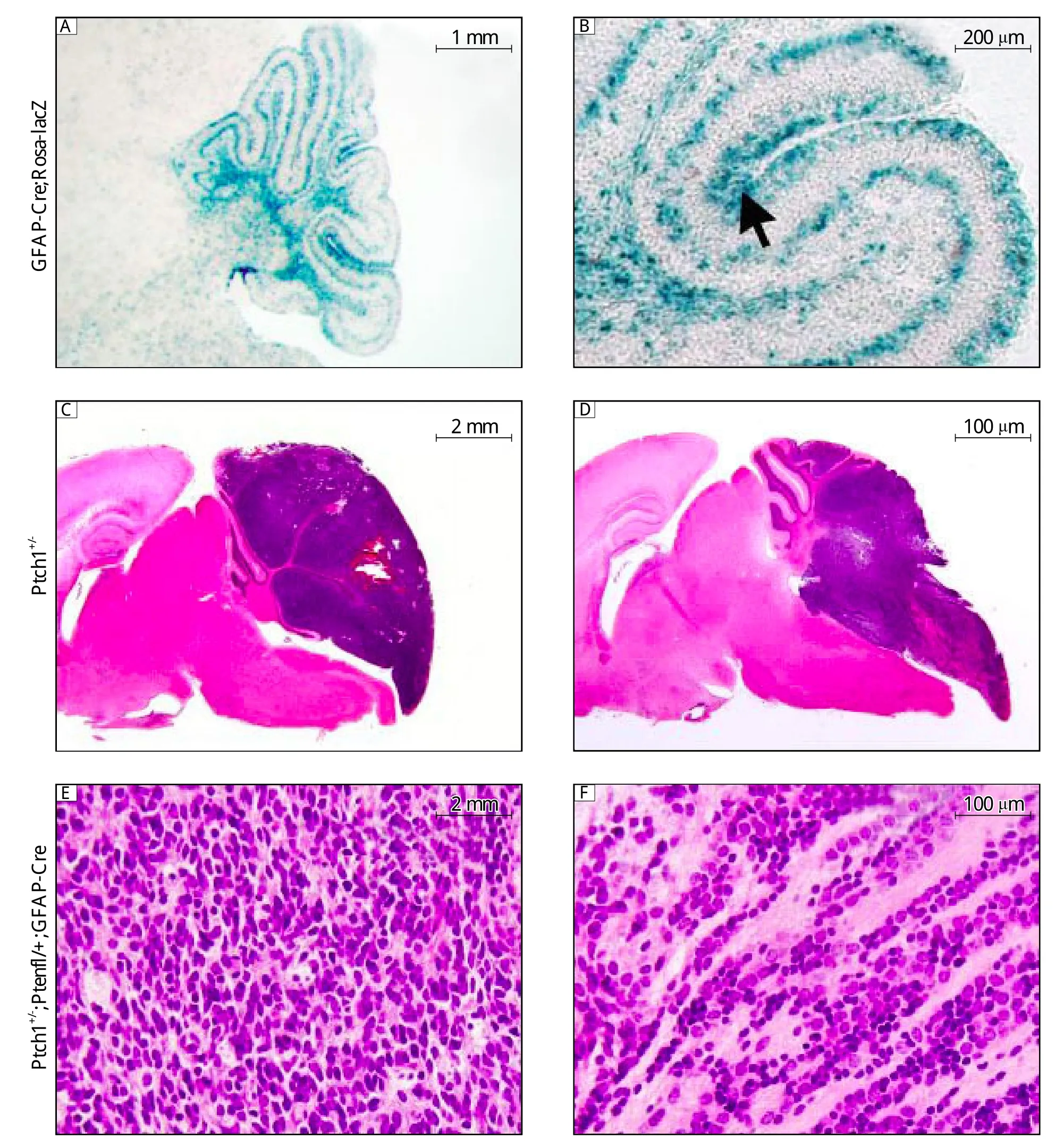
Fig. 1 Heterozygosity for Pten promotes medulloblastoma in the background of Ptch1+/-. A: β-galactosidase staining in GFAP-Cre;Rosa26-lacZ mice of P7. B: Higher magnification of (A). Positive for β-galactosidase staining (blue reaction product)was found both in astrocytes and in immature precursor cells of the external granular layer (arrow)in the developing cerebellum. C: H&E staining of histological sections of representative malignancies in Ptch1+/- medulloblastomas. D: H&E staining of histological sections of representative malignancies in Ptch1+/-; PtenFloxp/+;GFAP-Cre medulloblastomas. E: Higher magnification of (C). F: Higher magnification of (D).
Mouse medulloblastoma with heterozygosity for Pten and Ptch1 overexpresses microRNAs from the miR-183~96~182 cluster
The miR-183~96~182 cluster is a highly conserved cluster of microRNAs residing on an intergenic region of human chromosome 7q, which is commonly gained in medulloblastoma. The miR-183~96~182 cluster has been largely reported as pro-tumorigenic/prometastatic effects in cancers. Increased expression of the miR-183~96~182 cluster of microRNAs has been reported in several subgroups of medulloblastoma, including the most clinically aggressive subgroup associated with genetic amplification of MYC and activation of photoreceptor transcriptional programs[11,12]. Therefore, we investigated the expression of mmu-miR-183, mmumiR-96 and mmu-miR-182 in these medulloblastomas.The genes for 3 microRNAs of the cluster, including miR-183, miR-96 and miR-182, were clustered within 4 kb on mouse chr6qA3, and transcribed in the same direction (telomere→centromere)(Fig. 2A). All the 3 microRNAs had a high degree of sequence homology and potentially overlapping mRNA targets (Fig.2B). For the stem-loop RT-PCR assay, we found that the mmu-miR-183, mmu-miR-96 and mmu-miR-182 expressions were increased in Ptch1+/-;PtenFloxp/+GFAP-Cre medulloblastoma compared with Ptch1+/-medulloblastoma, especially the mmu-miR-183 and mmu-miR-96. However, as the same PCR cycles,the expression of these microRNAs had very low level in adult normal cerebellum of WT, GFAP-Cre,PtenFloxp/+;GFAP-Cre, Ptch1+/-and Ptch1+/-Pten-Floxp/+GFAP-Cre mice (Fig. 2C). Therefore, we speculated that high expression of the miR-183~96~182 cluster was an important factor in promoting tumorigenesis in Ptch1+/-PtenFloxp/+GFAP-Cre mice.
Enforced expression of miR-183~96~182 promotes CGNP proliferation in cooperation with the hedgehog signaling pathway
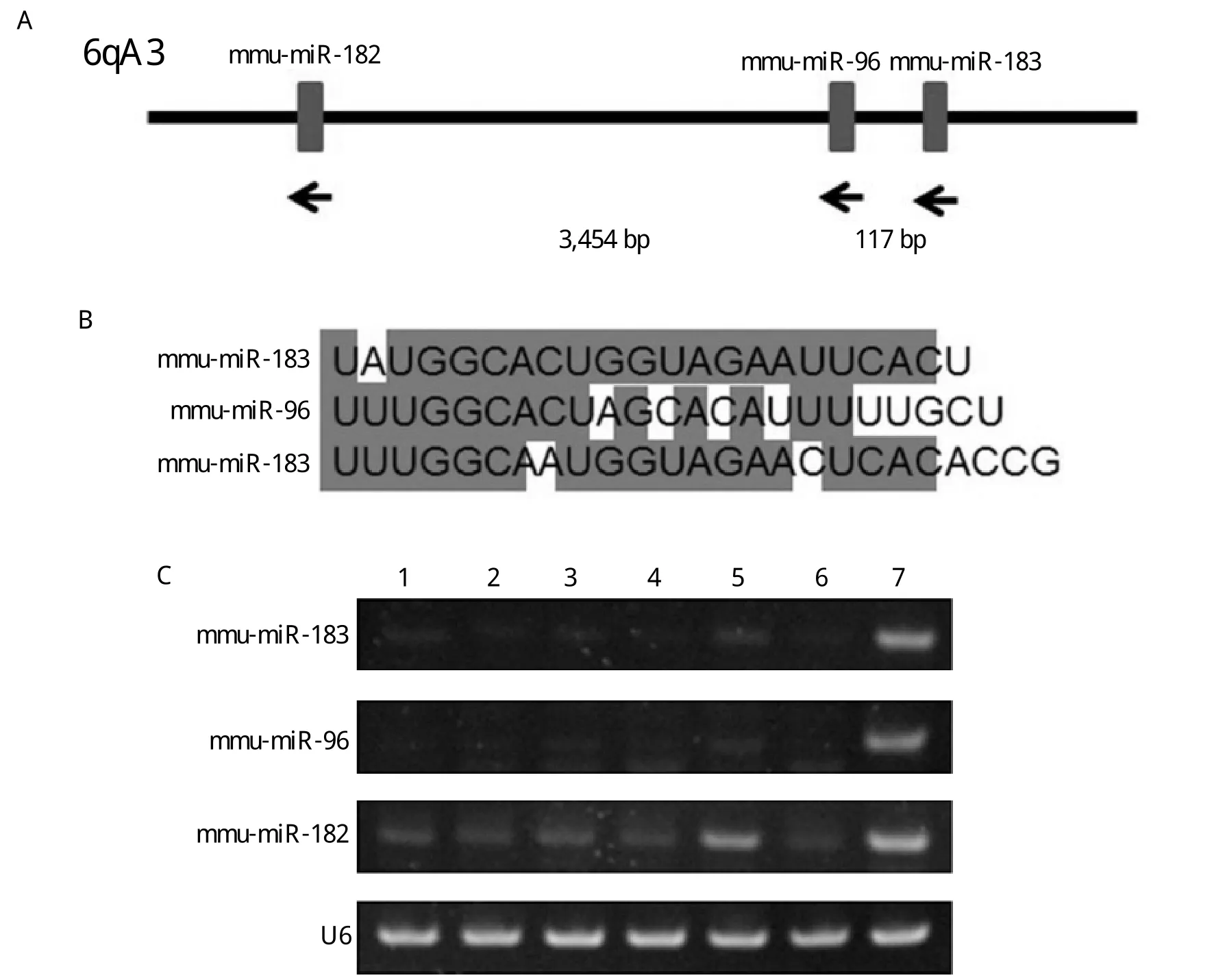
Fig. 2 Overexpresses miRNAs from the miR-183~96~182 Cluster in Ptch1+/-; PtenFloxp/+; GFAP-Cre medulloblastomas. A: Genomic organization of the miR-183~96~182 cluster on mouse chr6qA3. The numbers show the spacing in each of the three species in genes. B:Sequence alignment for mouse miR-183, miR-96, and miR-182. C: RT-PCR detection of mature miR-183, miR-96, and miR-182 in adult normal cerebellum of WT, GFAP-Cre, PtenFloxp/+;GFAP-Cre, Ptch1+/- and Ptch1+/-;PtenFloxp/+; GFAP-Cre mouse and medulloblastoma of Ptch1+/- and Ptch1+/;PtenFloxp/+;GFAP-Cre mouse. The U6 RNA was used for normalization. Lane 1: WT cerebellum; Lane 2: GFAP-Cre cerebellum; Lane 3: Ptenfl/+;GFAP-Cre cerebellum; Lane 4: Ptch1+/- cerebellum; Lane 5: Ptch1+/- medullobastoma; Lane 6: Ptch1+/- Ptenfl/+; GFAP-Cre cerebellum; Lane 7:Ptch1+/- Ptenfl/+; GFAP-Cre medulloblastoma.
As CGNPs are the source of Shh-subtype medulloblstomas, we asked that whether the overexpression of miR-183~96~182 promoted CGNP proliferation,and dysregulation of miR-183~96~182 was one of the most important factors in Ptch1+/-PtenFloxp/+GFAPCre medulloblastomas. We isolated CGNPs from the cerebellum of 7-day-old C57BL/6 pups and infected them with retroviruses expressing the mmumiR-183~96~182 cluster or control with Retro-GFP for 12 hours and then cultured CGNPs with or without ShhN CM. After infecting retrovirus for 36, 48, 60 and 72 hours, we measured each CGNP proliferation by quantifying EdU staining, respectively (Fig. 3A).Consistent with previous studies, we found that cells maintained proliferation in the presence of shhN. After infection for 36 hours, the overexpression of miR-183~96~182 had little effect on high concentration of ShhN. During 48 to 60 hours, the overexpression of miR-183~96~182 cluster increased proliferation in ShhN CM-treated cells (Fig. 3B). With the prolongation of the culture time, CGNPs grew with contact inhibition. The proliferation was reduced in the group of overexpressing miR-183~96~182 with ShhN CM at 72 hours. However, when cells were cultured in the absence of ShhN, they showed a decline in proliferation. In the beginning, the proliferation of miR-183~96~182-infected CGNPs was decreased slower than control at 36 hours, which then showed a continuous decline. There were no significant differences for proliferation between miR-183~96~182-infected CGNPs and control-infected CGNPs from 48 hours(Fig. 3C). Therefore, miR-183~96~182 was unable to maintain cell proliferation in the absence of Shh, suggesting that its role in promoting CGNPs proliferation is dependent on hedgehog signaling activation.
DISCUSSION
Chromosome 10q deletion is frequently identified in human medulloblastoma, and is shown to be highly associated with survival probability. Chromosome 10q loss was primarily limited to Shh and group C tumors[26]. Adult Shh-medulloblastomas harboring 10q loss show particularly poor progression-free survival and overall survival probabilities[27]. PTEN, the tumor suppressor gene of phosphatase and tensin homolog,is located on the chromosomal region 10q23.31. We established a mouse model to present the important role for cross talk between Pten loss and sonic hedgehog signaling pathways in the pathogenesis of medulloblastoma.
We inactivated Pten during brain development in neuronal and glial cell populations in a precisely defined area of the cerebellum. It was achieved by using GFAP-Cre transgenic mice, in which Cre is active in EGL, the Bergmann glia and white matter of the cerebellum. PtenFloxp/FloxpGFAP-Cre mice developed seizures and ataxia early in life and died prematurely,which is similar to other previous studies[28,29]. Although PTEN is frequently inactivated in malignant human brain tumors, inactivation of Pten in the mouse brain does not lead to tumor development, which may due to the fact that additional genetic alterations are necessary. We used the Ptch1-lacZ mouse model of medulloblastoma to study the effect of Pten loss on medulloblastoma tumorigenesis. We found that the heterozygosity for Pten, in the context of heterozygosity for Ptch1, altered tumor histology and accelerated medulloblastoma tumorigenesis in Ptch1+/-PtenFloxp/+GFAP-Cre mice. Although spontaneous medulloblastoma harbor only monoallelic mutations of Pten possess at least one functional wildtype Pten allele, they further or completely lose Pten protein immunoreactivity[30]. These results showed that epigenetic silencing by aberrant promoter methylation,deregulated transcription and/or increased degradation may disrupt function and promote tumorigenesis. As a result, loss of Pten expression leads to activation of PI3K signaling: increased expression of activated Akt,phosphorylated on serine 473, in Ptch1+/-PtenFloxp/+GFAP-Cre medulloblastoma (data not shown).
Here we examined the role of miR-183~96~182 in the context of heterozygosity for Pten and Ptch1 associated medulloblastoma. The miR-183~96~182 cluster that upregulated across Ptch1+/-PtenFloxp/+GFAP-Cre medulloblastoma samples is relative to Ptch1+/-medulloblastoma. MiR-183~96~182 has been implicated in retinal development and stem-cell maintenance[31-33]. With the expression of GFAP-Cre and recombination occur in scattered EGL cells before neoplastic transformation, we concluded that in Ptch1+/-PtenFloxp/+GFAP-Cre mouse model, medulloblastomas arise from cerebellar granule neuron precursors retained on the cerebellar surface. CGNPs taken from the P6~7 cerebellums have greater proliferative capacity than those harvested later, when neuronal progenitors have already begun to exit the cell division cycle and migrate into the internal granular layer.When explanted into culture in the absence of ShhN,CGNP rapidly exit the cell cycle and differentiate,but ShhN additionally extends their proliferative potential[19]. CGNPs harvested from C57BL/6 mice and engineered to ectopically express miR-183~96~182 by retroviral transfer demonstrated a greater proliferative advantage than their Retro-GFP counterparts in the presence of ShhN. However, its role in promoting CGNPs proliferation is dependent on hedgehog signaling activation. When cultured in vitro for 48 hours,CGNPs without ShhN-CM lost the effort of endogenous ShhN and declined sharply in the proliferation,despite enforced expression of miR-183~96~182. Our data concluded that miR-183~96~182 was unable to maintain cell proliferation in the absence of ShhN.miR-183~96~182 could amplify the effects of Shh signaling, perhaps by inhibiting a transcriptional repressor of a subset of Shh signaling target genes.
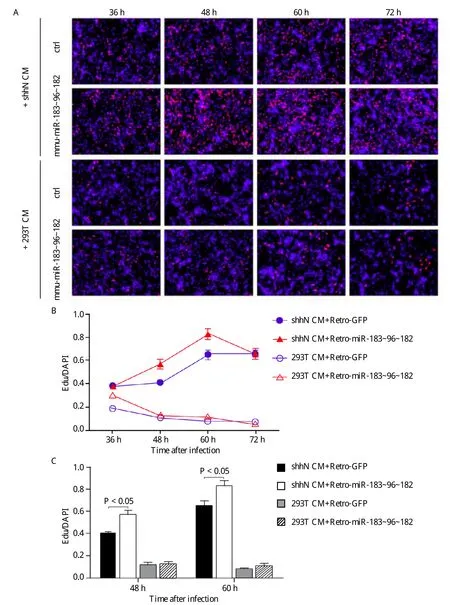
Fig. 3 Enforced expression of the miR-183~96~182 cluster promotes CGNPs proliferation. A: Immunofluorescence staining for EdU incorporation. Freshly isolated CGNPs were grown in poly-D-lysine and matrigel coated chamber slides in the presence of ShhN CM or 293T CM.The cells were infected with retroviral miR-183~96~182 or empty-control. B: Kinetics of proliferative response to miR-183~96~182 or empty-control in the presence ShhN CM or 293T CM. The numbers at each data point were counted from 4 random fields. C: Column diagram shows quantification of EdU-positive CGNPs after retroviral infection for 48 hours and 60 hours. The bars denote standard deviation.
In conclusion, our study provides evidence for an important role of miR-183~96~182 cluster in heterozygosity for Pten and Ptch1 associated medulloblastomas. Antagomirs to the miR-183~96~182 cluster might provide a potential new therapeutic strategy for patients with medulloblastomas harboring a constitutively activated PI3K signaling pathway.
1] Ellison D. Classifying the medulloblastoma: insights from morphology and molecular genetics. Neuropathol Appl Neurobiol 2002; 28: 257-82.
[2]Packer RJ, Cogen P, Vezina G, Rorke LB. Medulloblastoma: clinical and biologic aspects. Neuro Oncol 1999; 1:232-50.
[3]Heikens J, Michiels EM, Behrendt H, Endert E, Bakker PJ, Fliers E. Long-term neuro-endocrine sequelae after treatment for childhood medulloblastoma. Eur J Cancer 1998; 34: 1592-7.
[4]Mulhern RK, Palmer SL, Merchant TE, Wallace D, Kocak M, Brouwers P, et al. Neurocognitive consequences of risk-adapted therapy for childhood medulloblastoma.J Clin Oncol 2005; 23: 5511-9.
[5]Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer 2006; 6: 857-66.
[6]He L, Thomson JM, Hemann MT, Hernando-Monge E,Mu D, Goodson S, et al. A microRNA polycistron as a potential human oncogene. Nature 2005; 435: 828-33.
[7]Carmell MA, Hannon GJ. RNase III enzymes and the initiation of gene silencing. Nat Struct Mol Biol 2004;11: 214-8.
[8]Northcott PA, Fernandez LA, Hagan JP, Ellison DW,Grajkowska W, Gillespie Y, et al. The miR-17/92 polycistron is up-regulated in sonic hedgehog-driven medulloblastomas and induced by N-myc in sonic hedgehogtreated cerebellar neural precursors. Cancer Res 2009;69: 3249-55.
[9]Uziel T, Karginov FV, Xie S, Parker JS, Wang YD,Gajjar A, et al. The miR-17~92 cluster collaborates with the Sonic Hedgehog pathway in medulloblastoma. Proc Natl Acad Sci U S A 2009; 106: 2812-7.
[10]Cho YJ, Tsherniak A, Tamayo P, Santagata S, Ligon A, Greulich H, et al. Integrative genomic analysis of medulloblastoma identifies a molecular subgroup that drives poor clinical outcome. J Clin Oncol 2011; 29:1424-30.
[11]Bai AH, Milde T, Remke M, Rolli CG, Hielscher T,Cho YJ, et al. MicroRNA-182 promotes leptomeningeal spread of non-sonic hedgehog-medulloblastoma. Acta Neuropathol 2012; 123: 529-38.
[12]Weeraratne SD, Amani V, Teider N, Pierre-Francois J,Winter D, Kye MJ, et al. Pleiotropic effects of miR-183~96~182 converge to regulate cell survival, proliferation and migration in medulloblastoma. Acta Neuropathol 2012; 123: 539-52.
[13]Yang ZJ, Ellis T, Markant SL, Read TA, Kessler JD,Bourboulas M, et al. Medulloblastoma can be initiated by deletion of Patched in lineage-restricted progenitors or stem cells. Cancer Cell 2008; 14: 135-45.
[14]Rubin JB, Rowitch DH. Medulloblastoma: a problem of developmental biology. Cancer Cell 2002; 2: 7-8.
[15]Goodrich LV, Milenkovic L, Higgins KM, Scott MP.Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997; 277: 1109-13.
[16]Suzuki A, Yamaguchi MT, Ohteki T, Sasaki T, Kaisho T,Kimura Y, et al. T cell-specific loss of Pten leads to defects in central and peripheral tolerance. Immunity 2001;14: 523-34.
[17]Peng YJ, He WQ, Tang J, Tao T, Chen C, Gao YQ, et al.Trio is a key guanine nucleotide exchange factor coordinating regulation of the migration and morphogenesis of granule cells in the developing cerebellum. J Biol Chem 2010; 285: 24834-44.
[18]Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH,Nguyen JT, et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res 2005; 33:e179.
[19]Wechsler-Reya RJ, Scott MP. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron 1999; 22: 103-14.
[20]Zindy F, Uziel T, Ayrault O, Calabrese C, Valentine M,Rehg JE, et al. Genetic alterations in mouse medulloblastomas and generation of tumors de novo from primary cerebellar granule neuron precursors. Cancer Res 2007;67: 2676-84.
[21]Yue S, Chen Y, Cheng SY. Hedgehog signaling promotes the degradation of tumor suppressor Sufu through the ubiquitin-proteasome pathway. Oncogene 2009; 28:492-9.
[22]Read TA, Fogarty MP, Markant SL, McLendon RE, Wei Z, Ellison DW, et al. Identification of CD15 as a marker for tumor-propagating cells in a mouse model of medulloblastoma. Cancer Cell 2009; 15: 135-47.
[23]Uziel T, Zindy F, Xie S, Lee Y, Forget A, Magdaleno S,et al. The tumor suppressors Ink4c and p53 collaborate independently with Patched to suppress medulloblastoma formation. Genes Dev 2005; 19: 2656-67.
[24]Schuller U, Heine VM, Mao J, Kho AT, Dillon AK,Han YG, et al. Acquisition of granule neuron precursor identity is a critical determinant of progenitor cell competence to form Shh-induced medulloblastoma. Cancer Cell 2008; 14: 123-34.
[25]Gibson P, Tong Y, Robinson G, Thompson MC, Currle DS, Eden C, et al. Subtypes of medulloblastoma have distinct developmental origins. Nature 2010; 468: 1095-9.
[26]Northcott PA, Korshunov A, Witt H, Hielscher T, Eber-hart CG, Mack S, et al. Medulloblastoma comprises four distinct molecular variants. J Clin Oncol 2011; 29: 1408-14.
[27]Northcott PA, Hielscher T, Dubuc A, Mack S, Shih D,Remke M, et al. Pediatric and adult sonic hedgehog medulloblastomas are clinically and molecularly distinct.Acta Neuropathol 2011; 122: 231-40.
[28]Backman SA, Stambolic V, Suzuki A, Haight J, Elia A,Pretorius J, et al. Deletion of Pten in mouse brain causes seizures, ataxia and defects in soma size resembling Lhermitte-Duclos disease. Nat Genet 2001; 29: 396-403.
[29]Kwon CH, Zhu X, Zhang J, Knoop LL, Tharp R, Smeyne RJ, et al. Pten regulates neuronal soma size: a mouse model of Lhermitte-Duclos disease. Nat Genet 2001; 29:404-11.
[30]Castellino RC, Barwick BG, Schniederjan M, Buss MC,Becher O, Hambardzumyan D, et al. Heterozygosity for Pten promotes tumorigenesis in a mouse model of medulloblastoma. PLoS One 2010; 5: e10849.
[31]Loscher CJ, Hokamp K, Kenna PF, Ivens AC, Humphries P, Palfi A, et al. Altered retinal microRNA expression profile in a mouse model of retinitis pigmentosa. Genome Biol 2007; 8: R248.
[32]Loscher CJ, Hokamp K, Wilson JH, Li T, Humphries P, Farrar GJ, et al. A common microRNA signature in mouse models of retinal degeneration. Exp Eye Res 2008; 87: 529-34.
[33]Viswanathan SR, Mermel CH, Lu J, Lu CW, Golub TR,Daley GQ. microRNA expression during trophectoderm specification. PLoS One 2009; 4: e6143.
 THE JOURNAL OF BIOMEDICAL RESEARCH2013年6期
THE JOURNAL OF BIOMEDICAL RESEARCH2013年6期
- THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Multimodality image fusion for diagnosing coronary artery disease
- Human blood plasma-based electronic integrated circuit amplifier configuration
- Pulmonary cystic disease associated with integumentary and renal manifestations
- Volume conduction energy transfer for implantable devices
- Individual differences in transcranial electrical stimulation current density
- Hypoxia-induced factor-1 alpha upregulates vascular endothelial growth factor C to promote lymphangiogenesis and angiogenesis in breast cancer patients