
Production traits and fertility of reciprocal hybrids between Argopecten irradians irradians and A.i.concentricus*
2024-02-27 08:28:24JinjingWANGJunhaoNINGXiaLUMinCHENGuilongLIUXinXUQuanchaoWANGHeXUChundeWANG
Jinjing WANG, Junhao NING, Xia LU, Min CHEN, Guilong LIU, Xin XU,Quanchao WANG, He XU, Chunde WANG,2
1 Research and Development Center for Efficient Utilization of Coastal Bioresources, Yantai Institute of Coastal Zone Research,Chinese Academy of Sciences, Yantai 264003, China
2 College of Marine Science and Engineering, Qingdao Agricultural University, Qingdao 266109, China
3 University of Chinese Academy of Sciences, Beijing 100049, China
4 Yantai Spring-Sea AquaSeed Co., Ltd., Yantai 264006, China
5 Jiangsu Baoyuan Biotechnology Co., Ltd., Lianyungang 222144, China
6 Jiangsu Haitai MariTech Co., Ltd., Lianyungang 222144, China
Abstract The two bay scallop subspecies, Argopecten irradians irradians (NN) and A.i.concentricus(SS), are fast growing and major cultured bivalves in China.However, their relatively small sizes and decreasing production traits caused by long-term inbreeding have been major concerns to the industry in the last two decades.Hybridization between the two bay scallop subspecies may provide a new approach to breed a new variety with superior production traits for the industry.For this end, in this study, we hybridized the two bay scallop subspecies in order to obtain a new strain that incorporates the genes of both subspecies.No significant difference was found in fertilization rate, hatching rate and metamorphosis rate between the purebred and crossbred cohorts (NN♀×SS♂, denoted as NS; SS♀×NN♂, denoted as SN).Both mating strategy (intra- vs.inter-population crosses) and egg origin had significant effects on growth and survival at the larval stage.Heterosis was observed in the crossbred and was more pronounced in older stages.Genetic diversity of the reciprocal hybrids, especially that of SN, was increased compared with the purebred cohorts.Almost all hybrids were completely fertile and able to reproduce by selffertilization or by backcrossing with either parent.Apparently, male sterile individuals whose gonads were fully occupied by the ovary part at mature stage were found in the hybrids for the first time.The hybrids,especially SN, may provide precious germplasm resources for the production of ternary hybrids with the Peruvian scallop, A.purpuratus.
Keyword: bay scallop; hybridization; growth; survival; heterosis; fertility; ternary hybrids
1 INTRODUCTION
The two bay scallop subspecies,Argopecten irradiansirradians(NN) andA.i.concentricus(SS),were introduced to China from the United States in early 1980s and 1990s, respectively.With the advances of artificial breeding technology, bay scallop farming soon developed into a large industry along China coasts (Zhang et al., 1986, 2000).AlthoughArgopectenscallops are fast-growing bivalves, the two bay scallop subspecies are only about 5-6 cm in market size.And long-term inbreeding from limited number of originally introduced scallops has caused severe problems,such as poor growth, high mortality and economic losses to the industry (Zheng et al., 2007, 2008).Despite enormous efforts including re-introductions and intra-specific hybridization, the above problems remain largely unsolved (Zhang et al., 2007; Wang and Li, 2010; Wang et al., 2017).
Crossbreeding is a useful tool in genetic improvement of aquaculture species, which can effectively improve production traits and stress resistance of hybrid offspring (Zhang et al., 2007;Ahmed et al., 2008; Wang et al., 2011, 2017).To date, intra-specific and inter-specific hybridization have been successfully applied in various mollusks,including scallops (Yang et al., 2004; Zhang et al.,2007; Wang et al., 2011; Chen et al., 2020), oysters(Huo et al., 2013; Zhang et al., 2014; Yan et al.,2018) and abalones (Bao et al., 2002; Ahmed et al.,2008), resulting in several new molluscan strains with excellent traits.InArgopectenscallops, the medium-sized Peruvian scallop,A.purpuratus, was successfully introduced and hybridized with the two bay scallop subspecies, body weight of the reciprocal inter-specific hybrid F1cohorts was about 100% higher than the bay scallops parent (Wang et al., 2011, 2017).Three new strains, including“Bohai Red”, “QN-2,” and “QN Orange”, were selected from the F1hybrids betweenA.purpuratusandA.i.irradians(Wang et al., 2017; Xu et al.,2019; Chen et al., 2020).
Although body weight of these strains were increased by 38%-45% compared with the control bay scallops, their production traits including growth and survival were still much lower than those of the F1hybrids produced by the binary hybridization between the Peruvian scallops and either of the two bay scallop subspecies.The two bay scallop subspecies have distinct but complementary traits, such as larger shell size and cold tolerance for NN, while larger shell width and thermal resistance for SS.Besides, they have same number of chromosomes and similar karyotypes.Therefore, it is possible to obtain a superior strain incorporating the good traits of both subspecies,which used for future production of ternary hybrid scallops with the Peruvian scallop.The aim of this study was to establish and evaluate the performance of the reciprocal hybrids between NNand SS.
2 MATERIAL AND METHOD
2.1 Brood stock
In late December 2020, bay scallop northern subspeciesA.i.irradians(NN, with shell height of 5.5-6.9 cm) and southern subspeciesA.i.concentricus(SS, with shell height of 4.8-6.1 cm) adults were collected from a scallop farm located at Laizhou,Shandong Province, China and transported to the hatchery of Yantai Sea-Spring AquaSeed Co., Ltd.The SS cohort was the F2offspring of the southern population that was transported to Laizhou from Zhanjiang, Guangdong Province in spring 2018 and was reproduced with about 500 individuals in the hatchery for one generation each year.The two scallop stocks were maintained in sand-filtered seawater (with temperature of 10±0.5 °C, salinity of 27±1) for two weeks.After the acclimatization, the seawater temperature was manipulated from 10 °C gradually to 20 °C to ensure that both stocks matured at the same time.During the indoor conditioning period, scallops were fed with excessiveChaetocerosmuelleri,IsochrysisgalbanaandPlatymonassubcordiformisfour times a day and the feeding density was (2×104)-(3×104) cells/mL per day.In addition, the seawater was changed twice every day.
2.2 Spawning and fertilization
Mature brood stocks were induced to spawn separately by exposure to air for 30 min followed by a thermal shock from 20 to 24 °C.In brief, each scallop was placed in a 1-L beaker filled with 24-°C sand-filtered seawater to induce spawning, and closely watched to collect eggs or sperm separately.The gametes were collected immediately after spawning, and the beaker was then well washed with fresh water and filled with 24-°C seawater to collect more gametes.Eggs were checked under microscope for self-fertilization caused by sperm contamination and were discarded if they were found fertilized (Wang et al., 2011).The unfertilized eggs or sperm of each species from >100 scallops were pooled together and divided into two parts.Eggs from one species were mixed with sperm from the same species or another species to produce purebred or crossbred cohorts.Four crosses were produced, including NN♀×NN♂ (NN), SS♀×SS♂ (SS),NN♀×SS♂ (NS), and SS♀×NN♂ (SN), respectively.Three replicates were set up for each cross.The fertilized eggs were hatched in a 100-L plastic bucket at a density of 10-15 eggs/mL at 24 °C with aeration.Embryonic development in 24- °C seawater was observed consecutively under microscope.
2.3 Larval rearing, spat nursing, and grow-out
About 24 h after fertilization, D-larvae from each cohort (NN, SS, NS, and SN) were collected with a 50-μm sieve and cultivated in a 400-L bucket,separately.The culture density of larvae in the tank was maintained at 10-15 larvae/mL.The larvae were fed withI.galbanain the first 5 days and with a mixture ofI.galbanaandC.muelleri(2∶1 in volume) after day 5, in density of microalgae increased gradually from 20 000 to 80 000 cells/mL per day.The growth and survival of larvae in purebred and crossbred cohorts were examined continuously.
When about 40% of the larvae developed eyespots (at a shell length of 190-200 μm), plastic collectors were placed in the buckets for larva to set on.About 7 days after larval settlement, the collectors were transferred into a shrimp pond for spat nursing.About 6 weeks in pond, the spats were transferred into bags with larger mesh and cultured in open sea.The scallops were then placed in 10-layer lantern nets for adult grow-out in the sea when they exceeded 1.5 cm in shell height.The pure and hybrid cohorts were cultured in Yangma Island,Yantai.Thirty scallops from each cohort were randomly selected each month for measurements of their shell height, shell length, shell width, whole body weight, gonad weight, adductor muscle weight, and survival rate, respectively.
2.4 Histological observation
At grow-out stages from June to December 2021,ten scallops from each cohort were collected biweekly and dissected to collect their gonads.The dissected gonads were fixed in 4%paraformaldehyde at 4 °C for more than 24 h,dehydrated with series of ethanol (70%, 80%, 90%,95%, and 100%) and xylene, and then embedded in paraffin.The dehydrated gonads were serially sectioned at 5-μm sections and dewaxed with xylene after rehydration with gradient ethanol (100%, 95%,80%, and 70%), and finally stained with hematoxylin and eosin for observation (Ning et al.,2021).The developmental stages of gonads were observed under microscope and photographed by a high-resolution microscope camera.
2.5 Genetic diversity analysis
Genomic DNA was extracted from adult adductor muscles of the pure and hybrid cohorts using standard phenol/chloroform extraction method(Sambrook et al., 1989).The simple sequence repeats (SSR) primers used in this study were according to the published literatures (Table 1; Zhan et al., 2005; Li, 2009; Li et al., 2012).

Table 1 Primer sequences for the genetic diversity analysis
The PCR reactions were performed as follows:94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing for 1 min, and extension at 72 °C for 1 min, and finished with a final extension at 72 °C for 5 min.The PCR products were separated by electrophoresis on a 15% non-denaturing polyacrylamide gel, stained by silver staining and sized with the DL2000 Markers.
2.6 Statistical analysis
Fertilization, hatching, survival, and growth rates of the purebred and crossbred cohorts were statistically analyzed.To improve the normality and homogeneity of the data were tested and tenlogarithm or arcsine transformed if necessary.Differences in fertilization, hatching, survival and growth rates among the cohorts were analyzed with a one-way ANOVA (analysis of variance) followed by Duncan tests.Statistical analyses of the data were performed using the SPSS 26.0 software package and differences were considered statistically significant atP<0.05 (*) andP<0.01 (**).
Fertilization rate (%)=number of fertilized eggs/total number of eggs×100;
hatching rate (%) number of D-larvae/number of fertilized eggs×100;
metamorphosis rate (%)=number of metamorphosed larvae/number of larvae used×100;
survival rate (%)=number of surviving scallops/number of scallops used×100.
To determine the effects of egg origin (egg origin,the north scallop vs.the south scallop dams) and mating strategy (MS, intra- : NN♀×NN♂ and SS♀×SS♂ vs.inter- : NN♀×SS♂ and SS♀×NN♂population crosses) on growth and survival, a twofactor analysis of variance model was used according to previous studies (Cruz and Ibarra,1997; Zhang et al., 2007; Wang and Li, 2010; Wang et al., 2011; Wang and C?té, 2012; You et al., 2015):
whereYijkis the mean shell length (or survival rate)of thekreplicate from theiegg origin and thejmating strategy.EOiis the effect of egg origin on the shell length (survival rate) (i=1, 2).MSjis the effect of mating strategy on the shell height(survival rate) (j=1, 2).(EO×MS)ijis the effect of the interaction between the egg origin and the strategy, and eijkis the random observation error(k=1, 2, 3).
Heterosis was calculated using the method described in previous study (Cruz and Ibarra, 1997):Heterosis%=(F1-(NN+SS)/2)×100/((NN+SS)/2),where F1is the shell length (or survival rate) of reciprocal crosses while NN and SS are those of the purebred cohorts.
3 RESULT
3.1 Embryonic and larval development of the hybrids
The hybrid cohorts (NS and SN) were successfully produced by hybridizing NN and SS scallops and developed normally during embryonic and larval stages.As shown in Fig.1, the pure and hybrid cohorts underwent similar embryonic and larval development processes, including fertilization, cleavage, blastula,gastrula, trochophore larvae, D-larvae, eyespot larvae,settlement, and metamorphosis.
As depicted in Table 2, the fertilization rate in the SN cohort was significantly lower than NN but not different with other cohorts.No significant difference in hatching rate, initial D-larvae shell height, metamorphosis rate and development time was detected among these cohorts (Tables 2 & 3).
3.2 Heterosis in growth and survival at the larval and juvenile stage
The reciprocal hybrid cohorts (NS and SN) were significantly larger than the pure cohorts (NN and SS) at larval stage, except NS on Day 5.Heterosis in shell length of the hybrid cohorts increased gradually with growth and reached 17.01% in NS and 17.43% in SN on Day 30.On Days 5 and 20,the growth was significantly affected by egg origin and mating strategy, but not their interaction.On Days 10 and 25, the growth was only significantly affected by mating strategy.The growth on Days 15 and 30 was significantly affected by mating strategy,egg origin and their interaction (Tables 4 & 5).
3.3 Heterosis of growth and survival the pure and hybrid cohorts during grow-out stage
As shown in Table 6, NN cohort outperformed SS cohort in most time of the grow-out stage.Most importantly, the shell length and wet weight of hybrid cohorts (NS and SN) were significantly larger than those of pure cohorts (NN and SS)during the grow-out stage (P<0.05).The SS cohort was smaller than all the others in shell length were(P<0.05).
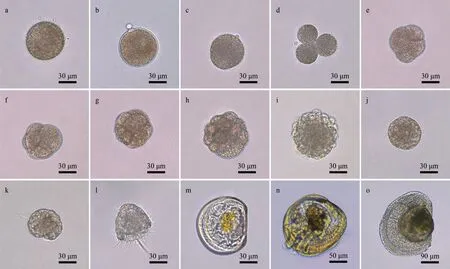
Fig.1 Embryonic and larval development of the purebred and crossbred cohorts

Table 2 Comparison of fertilization rate, hatching rate, initial D-larvae size, and metamorphosis rate among the pure and crossbred cohorts

Table 3 The development time of the four scallop cohorts at embryonic and larval stages

Table 4 Growth in shell length (μm) of the pure and hybrid cohorts in the larval and juvenile stages
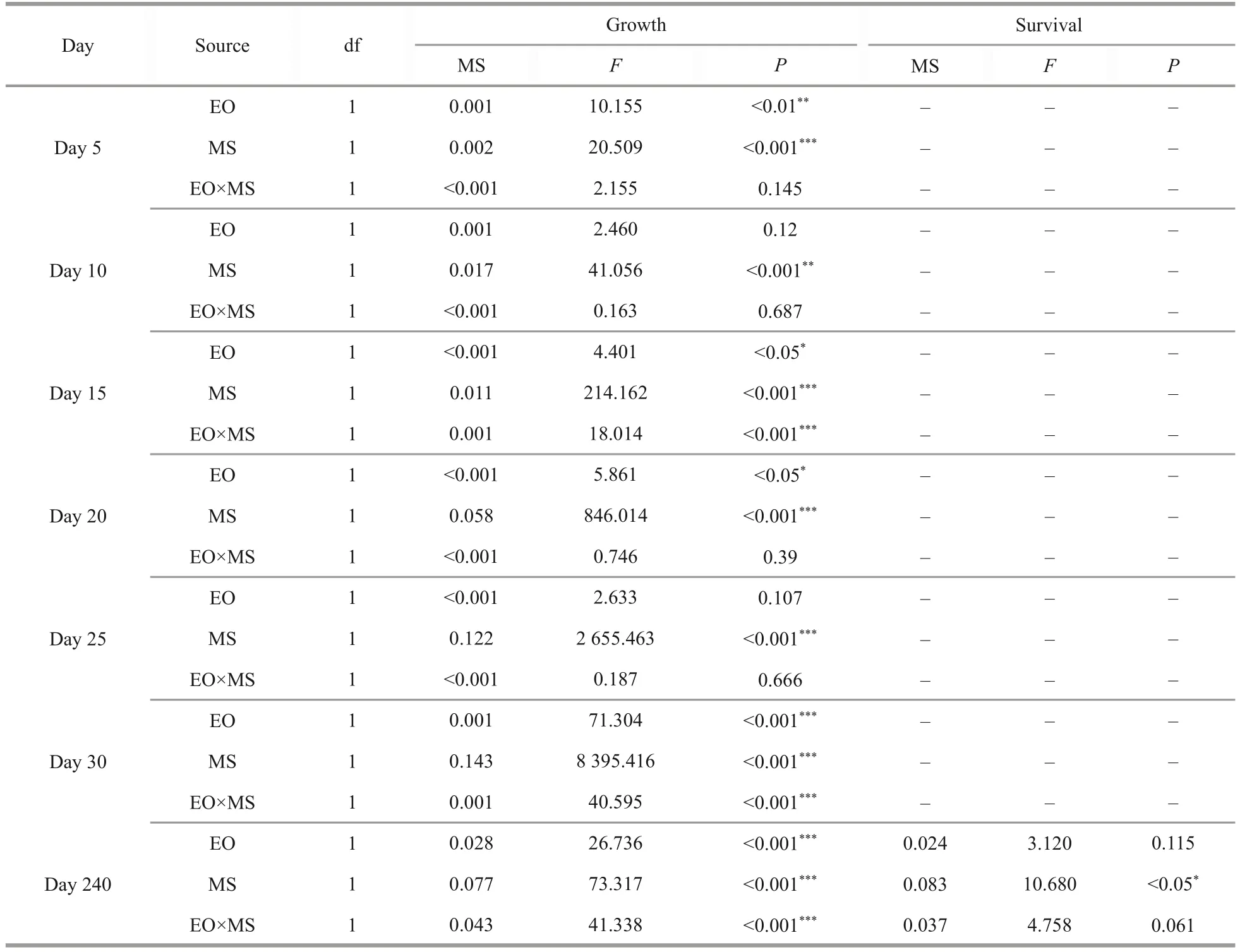
Table 5 Analyses of variance (ANOVA) of the effects of egg origin (EO) and mating strategy (MS) on growth and survival
At harvest on Day 240, the heterosis in shell length was 13.52% and 11.00% in NS and SN cohorts, respectively, while those in whole weight were 22.52% and 24.69% in NS and SN cohorts,respectively (Table 6).Both hybrid cohorts had larger shell width than the purebred cohorts (Fig.2).The SN cohort outperformed all other cohorts in the weight of adductor muscle, the main product ofArgopectenscallops.In terms of survival rate, both hybrid cohorts survived better than the purebred cohorts (Fig.2).On Day 240, the growth was significantly affected by egg origin, mating strategy and their interaction, and the survival rate was significantly affected by mating strategy only(Tables 5-6).
3.4 Genetic diversity analysis
Analyses of the genetic diversity utilizing 11microsatellite loci showed that the polymorphism level was significantly different between the pure and hybrid cohorts (Table 7).The number of alleles per locus ranged between 1.209 5 and 1.628 6, and that of the effective alleles varied from 1.154 0 to 1.468 2.The hybrid cohorts (especially SN) had more polymorphic loci and larger number of effective alleles than that in the pure cohorts, and theNa,Ne,Hgene diversity index and Shannon’s information index in hybrid cohorts were larger than those of the pure cohorts.
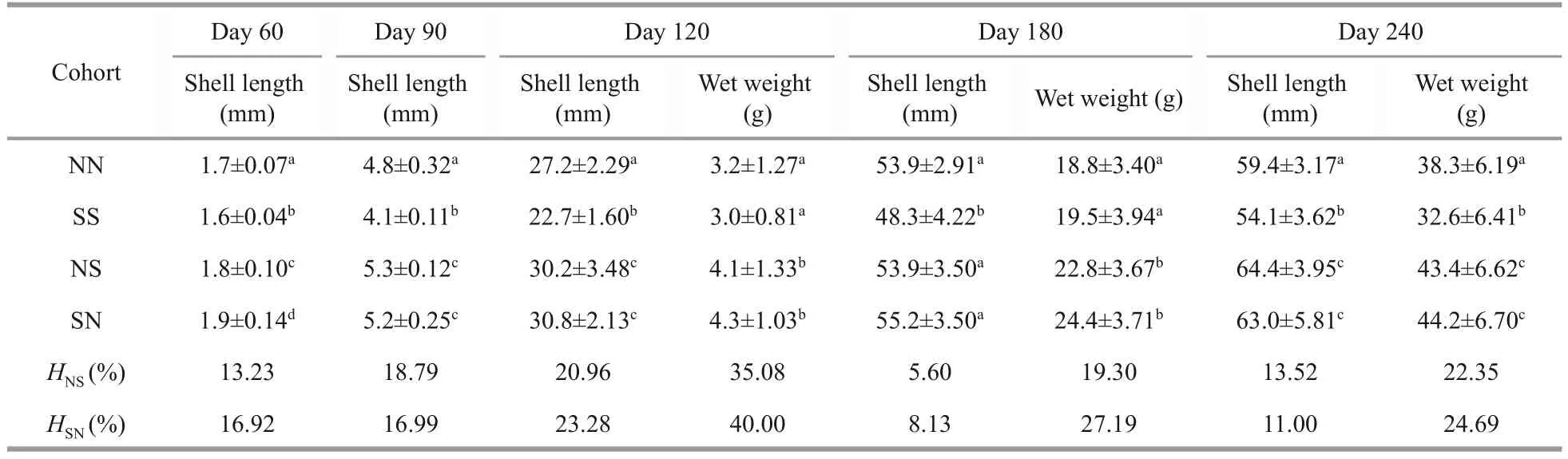
Table 6 Growth, survival, and the heterosis of hybrid and pure cohorts during grow-out stage

Fig.2 Sizes and survival of the pure and hybrid cohorts at harvest on Day 240
The genetic distance and genetic identity analyses showed that high identity and small genetic distance were observed between NN and NS as well as between SS and SN (Table 8).Additionally, a higher genetic identity in genotype was observed between NS and SN, while a lower genetic identity in genotype was detected between NN and SS.
3.5 Gonadal differentiation and fertility
Morphological and histological examinationsshowed that most scallops in the hybrid cohorts were hermaphroditic animals and their gonads developed normally as those of the purebred cohorts(Figs.3, 4a, & b).These hybrids were able to reproduce by self-fertilization or by crossing with either bay scallop subspecies.However, we found a few apparently dioecious females in the hybrid cohorts for the first time (Fig.4c-d).Unlike hermaphroditic scallops whose gonads have distinct whitish male and orange female part (Fig.4a-b), the gonads of these apparently dioecious females seemed to be occupied by only orange ovary, and no sperm or spermatids were found by histological examination.The gonad/somatic index (GSI) in the hybrid and purebred cohorts exhibited similar trends over the year (Fig.5).All cohorts experienced a decrease in GSI in September and October, followed by a gradual increase in the following months.
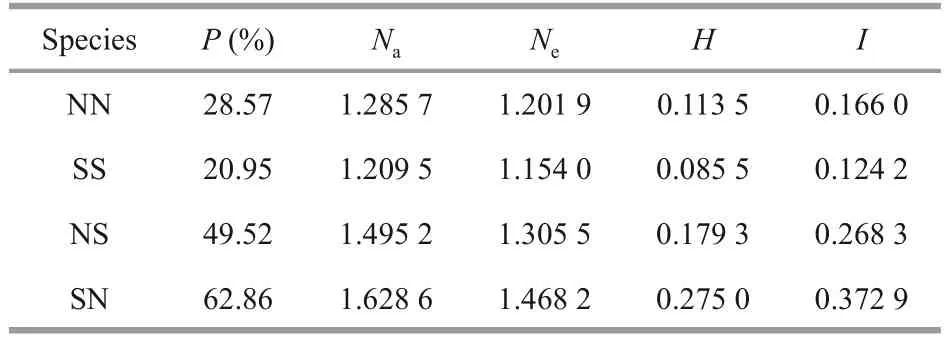
Table 7 Genetic diversity parameters of the hybrid and pure cohorts
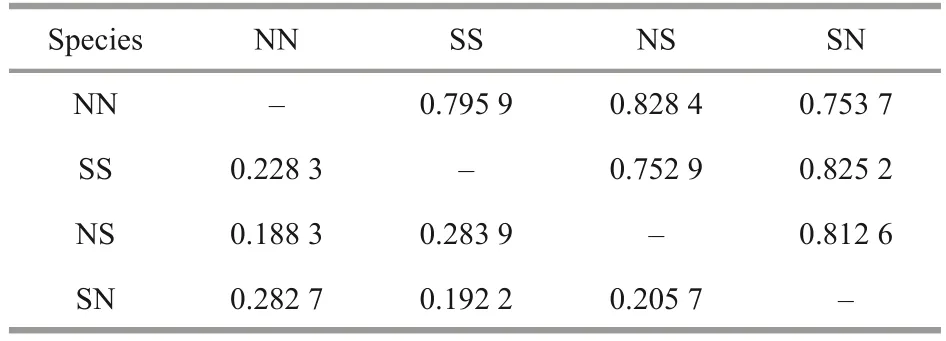
Table 8 Nei’s genetic identity and genetic distance of the hybrid and pure cohorts
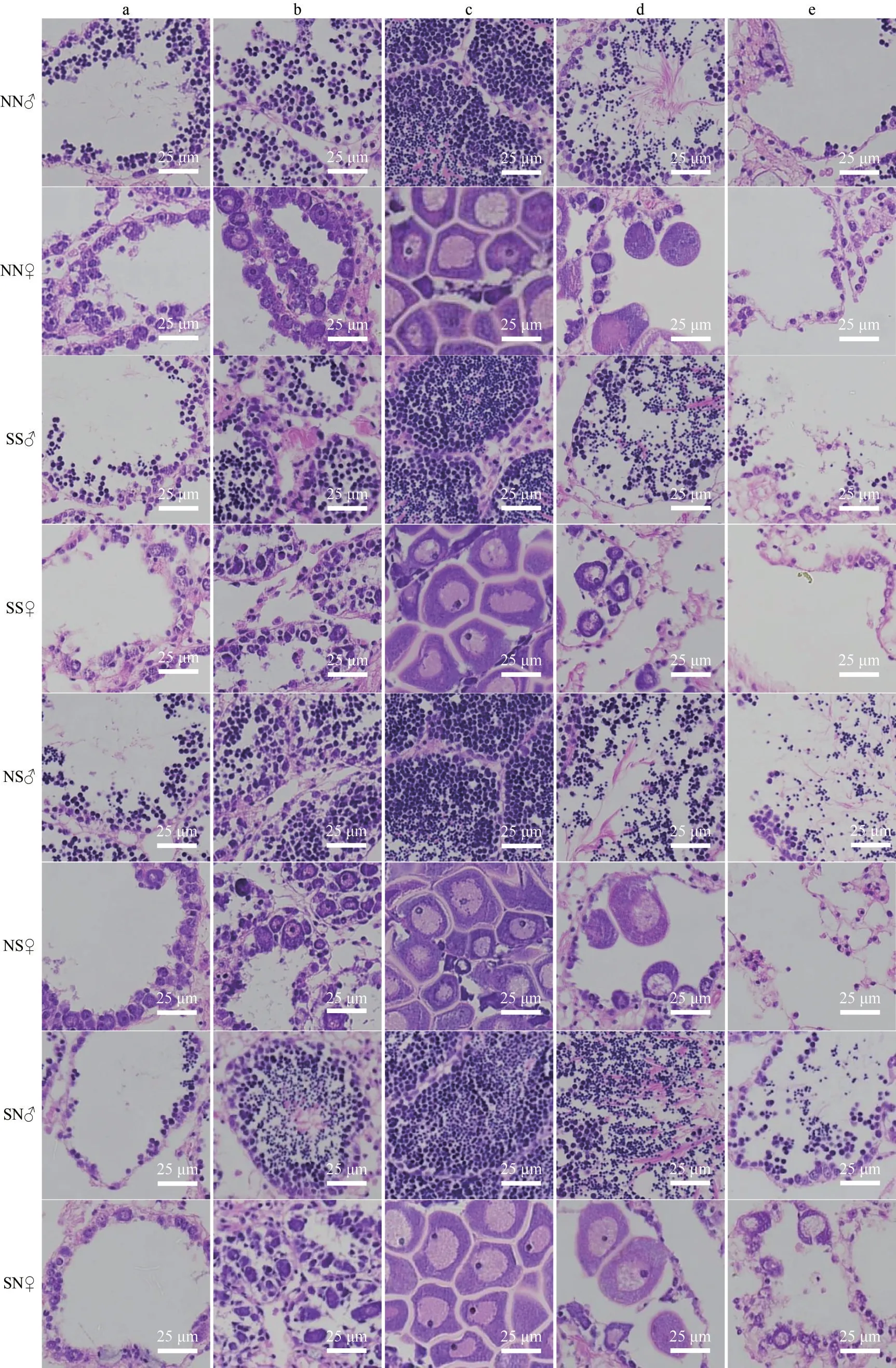
Fig.3 The gonadal development of the pure and hybrid cohorts
4 DISCUSSION
4.1 Growth and survival of the hybrid cohorts
In this study, both the SN and NS hybrid cohorts exhibited high fertilization and hatching rates,developed and grew normally from larval through to adult stage.Although no difference was detected in initial D-larvae size, the hatching duration of hybrid cohorts was significantly longer than purebred cohorts.The heterosis in growth and survival in hybrids (especially in the SN cohort) were significantly larger than that in pure lines, indicating that it is possible to improve the performance of the stocks by hybridizing the two subspecies.Similar results have been reported in hybridization ofArgopectenscallops (Cruz and Ibarra, 1997; Zhang et al., 2007; Wang et al., 2009, 2011).Hybridization of the bay scallop northern subspecies with Peruvian scallops resulted in F1hybrids with significantly elevated whole weight, up to about 100% higher than that of the control cohorts (Wang et al., 2009,2011).The success of hybridization betweenArgopectenscallops was mainly attributed to the fact that allArgopectenscallops have same chromosome number (2n=32) and similar karyotypes (5st+11t) (Wang and Guo, 2004; Zhang et al., 2007; Wang et al., 2011).Since the SN and NS hybrids outperformed their parents (NN and SS),we can expect that hybrids between SN or NS and the Peruvian scallops would also outperform those between SS or NN and the Peruvian scallops.
Hybridization can transfer excellent traits, such as growth, survival and stress tolerance of both parents to offspring, and even produce superior traits that are not possessed in one parent (Feng et al., 2012; Zhang et al., 2017; Tan et al., 2019; Qin et al., 2020; Xing et al., 2022).In this study, heterosis in growth and survival was found at both larval and adult stages.For instance, the wet weight and adductor muscle weight of hybrids were increased over 13% and 25% compared with the bay scallopA.i.irradians, and over 33% and 26% compared withA.i.concentricus, respectively.Similarly, the hybrids ofA.i.irradiansandA.purpuratusexhibited extraordinarily high heterosis in growth, and the production of hybrids can be increased by about 90% in whole body weight and about 150% in adductor muscle weight.The large difference in heterosis between the two studies may be caused by the fact that the hybridization between the bay scallop and Peruvian scallop is distant inter-specific hybridization while that between the two bay scallop subspecies are intra-specific hybridization.The wider range of genetic variations between two species than those between two subspecies may be the basis for the significant improvement in production traits in aquatic animals (Hu et al.,2013).In our study, the performance including growth and survival of SN was better than that of NS, making it the first choice for further hybridization with Peruvian scallops.
4.2 Genetic diversity of the hybrid cohorts

Fig.4 The hermaphroditic gonads of purebred (a) and hybrid (b) scallops and the dioecious gonads in the hybrid cohorts(c & d)
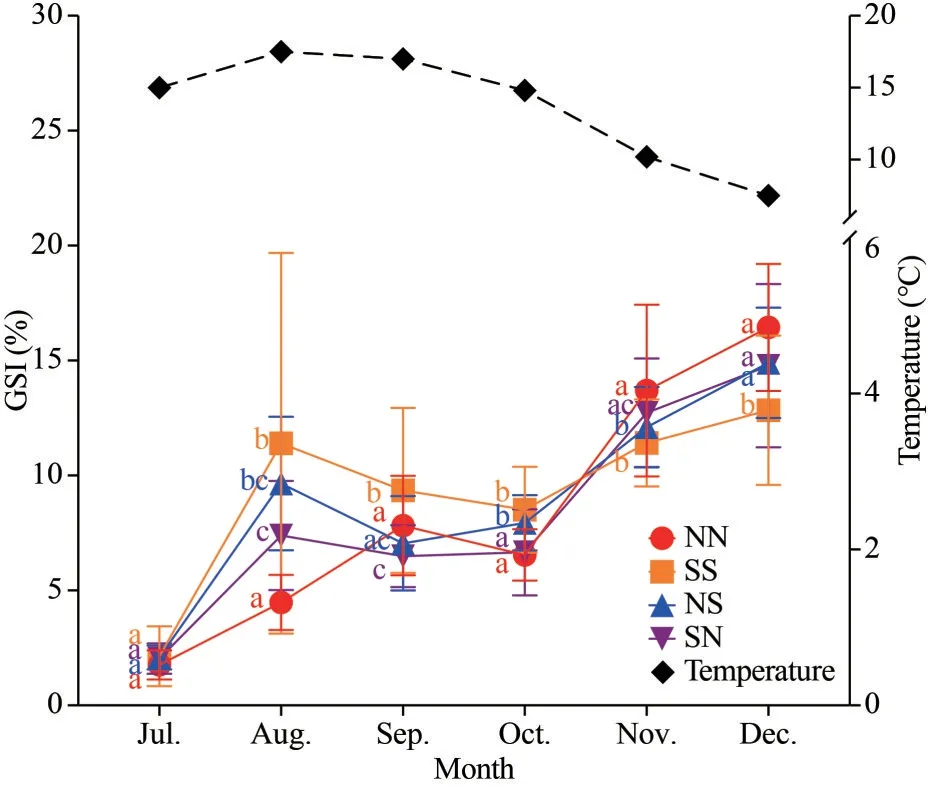
Fig.5 Annual change of GSI in the purebred and crossbred cohorts
Hybridization can also produce extraordinary heterosis in production traits by increasing the genetic diversity and gene heterozygosity in the hybrids (Li et al., 2011; Hu et al., 2013; Zhang et al.,2015).In this study, genetic diversity in hybrids was obviously higher than that in purebred cohorts.Meanwhile, the genetic diversity in SN cohort was higher than that of NS cohort, and thus produced larger heterosis in growth and survival.The high identity and small genetic distance observed between a hybrid cohort and its female parent (such as NN vs.NS and SS vs.SN) may be a direct result of the maternal effects.It is consistent with the results that gene loci of hybrid offspring were also genetically biased towards the female parent reported in the hybridization betweenChlamysfarreriandPatinopectenyessoensis(He et al.,2007).The fact that F1hybrids inherited nuclear material from both parents and the cytoplasmic genetic materials maternally may explain why traits and gene loci of hybrid offspring were more biased towards female parent (Hu et al., 2013).
4.3 Gonadal development and fertility of the hybrid cohorts
For the success of further hybridization of SN or NS hybrids with the Peruvian scallops, the SN and NS hybrids have to be male- and/or female-fertile.However, it has been recognized that inter-specific hybridization can lead to hybrid sterility or reduction of reproductive capacity owing to abnormality of meiosis and gonad development(Bartley et al., 2000).The typical example is that most of the hybrids betweenA.i.irradiansandA.purpuratuswere both male- and female-sterile and seemed to be morphologically and functionally females in which the ovary part occupied the whole gonad at mature stage (Wang et al., 2011, 2017).The phenomenon of hybrid sterility was also observed in inter-specific crossed oysters (Su et al.,2015; Zhang et al., 2017).Fortunately, in this study,all the reciprocal hybrids betweenA.i.irradiansandA.i.concentricuswere completely fertile and able to reproduce by self-fertilization or by backcrossing with either parent.Although fertility is common in intraspecific hybridization (Zhang et al., 2012,2016; Qin et al., 2020), we did find a few hybrids with apparently dioecious female gonads, as have been reported in the hybrids ofA.i.irradiansandA.purpuratus(Wang et al., 2009, 2011).The difference is that eggs of these hybrids in this study were completely fertile, which creates precious germplasm resources for further hybridization with the Peruvian scallop.
5 CONCLUSION
In conclusion, successful hybridization betweenA.i.irradiansandA.i.concentricuswas carried out in this study.Compared to pure scallop population,reciprocal hybrids exhibited high genetic diversity and significant heterosis in growth and survival.Almost all reciprocal hybrids were completely fertile, and their gonads developed normally.A few hybrids seemed to be morphologically and functionally females in which the ovary part occupied the whole gonads at mature stage.The NS and SN hybrid cohorts were precious germplasm resources for further hybridization with the Peruvian scallop.
6 DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
7 ACKNOWLEDGMENT
The authors are grateful to Min SHEN, Weian CAO, Qiang PENG, Zebin FENG, Fukai WANG,Xiaotong ZHANG and Kai YU, who come from Yantai Institute of Coastal Zone Research and Qingdao Agricultural University for their assistance during the hatchery, nursery and adult grow-out stages.
 Journal of Oceanology and Limnology2024年1期
Journal of Oceanology and Limnology2024年1期
- Journal of Oceanology and Limnology的其它文章
- Lagrangian coherent structure analysis on transport of Acetes chinensis along coast of Lianyungang, China*
- Transcriptome analysis reveals immune-related genes in tissues of Vibrio anguillarum-infected turbot Scophthalmus maximus*
- Effects of Poria cocos polysaccharide on growth performance, physiological parameters, and lipid metabolism of spotted sea bass Lateolabrax maculatus*
- A new diatom genus Spargeria gen.nov.(Bacillariophyceae)from Xizang, China, and the description of two new species*
- Lineaperpetua gen.nov.: a new diatom genus in the Thalassiosirales supported by morphology and molecular data*
- Neomicrocoleus gen.nov.(Microcoleaceae, Oscillatoriales), a novel cyanobacterial genus from benthic mats in a water channel*