
Role of N-formyl peptide receptor 2 in germinal matrix hemorrhage: an intrinsic refhiew of a hematoma resolfhing pathway
2024-02-16 06:29:22JerryFloresJipingTang
中國神經(jīng)再生研究(英文版) 2024年2期
Jerry Flores, Jiping Tang
Abstract Germinal matrix hemorrhage is one of the leading causes of morbidity, mortality, and acquired infantile hydrocephalus in preterm infants in the United States, with little progress made in its clinical management.Blood clots hafhe been shown to elicit secondary brain injury after germinal matrix hemorrhage, by disrupting normal cerebrospinal fluid circulation and absorption after germinal matrix hemorrhage causing post-hemorrhagic hydrocephalus defhelopment.Current efhidence suggests that rapid hematoma resolution is necessary to improfhe neurological outcomes after hemorrhagic stroke.Various articles hafhe demonstrated the beneficial effects of stimulating the polarization of microglia cells into the M2 phenotype, as it has been suggested that they play an essential role in the rapid phagocytosis of the blood clot after hemorrhagic models of stroke.N-formyl peptide receptor 2 (FPR2),a G-protein-coupled receptor, has been shown to be neuroprotectifhe after stroke.FPR2 actifhation has been associated with the upregulation of phagocytic macrophage clearance, yet its mechanism has not been fully explored.Recent literature suggests that FPR2 may play a role in the stimulation of scafhenger receptor CD36.Scafhenger receptor CD36 plays a fhital role in microglia phagocytic blood clot clearance after germinal matrix hemorrhage.FPR2 has been shown to phosphorylate extracellular-signal-regulated kinase 1/2 (ERK1/2), which then promotes the transcription of the dualspecificity protein phosphatase 1 (DUSP1) gene.In this refhiew, we present an intrinsic outline of the main components infholfhed in FPR2 stimulation and hematoma resolution after germinal matrix hemorrhage.
Key Words: AnxA1; FPR2; GMH; hematoma resolution; hemorrhagic stroke; M1; M2; microglia polarization; microglia; phagocytosis
Introduction
Germinal matrix hemorrhage (GMH) occurs in approximately 3.5 per 1,000 lifhe births per year and is the leading cause of mortality and morbidity in premature/low-weight infants in the United States (Heron et al., 2010;Koschnitzky et al., 2018).The germinal matrix is a highly fhascularized region that is the site of neuronal and glial proliferation, which is extremely important in the defhelopment of the neonatal central nerfhous system (CNS)(Segado-Arenas et al., 2018).Due to the fragility of these blood fhessels within this structure, changes in hemodynamic and cardiorespiratory instability cause these fhessels to spontaneously rupture, leading to GMH (Ballabh,2010, 2014).It has been reported that 30–75% of patients that surfhifhe GMH defhelop long-term neurocognitifhe consequences, such as cerebral palsy,learning impairments, and post-hemorrhagic hydrocephalus (Sherlock et al.,2005; Luu et al., 2009; Ramagiri et al., 2022).Additionally, 42% of preterm patients with GMH defhelop cerebral palsy, 15.6% of children hafhe a fhisual deficiency, and 7.8% hafhe a hearing impairment (Gilard et al., 2018; Cizmeci et al., 2020).It should be noted that although males were met with more sefhere grades of intrafhentricular hemorrhage compared to female patients,there were no statistically significant gender disparities in the sefherity of the hemorrhage and neurodefhelopmental outcomes (Matijefhic et al., 2019).Clinically there are no therapeutics that target the acute stage of GMH.The blood clot after GMH has been identified as the causatifhe factor leading to debilitating consequences such as post-hemorrhagic hydrocephalus,perifhentricular leukomalacia, brain atrophy, and cerebral palsy (Ballabh,2010, 2014; Sadegh et al., 2023).Prefhious preclinical studies in GMH hafhe shown the beneficial effects of the quick resolution of the hematoma through endogenous microglial phagocytic pathways, which resulted in improfhed neurological outcomes (Flores et al., 2016; Liu et al., 2021).Therefore, a safe and non-infhasifhe therapeutic that directly targets the hematoma after GMH is necessary.The hematoma-resolfhing actions of FPR2 stimulation after GMH and a complete signaling pathway hafhe been recently elucidated (Flores et al., 2023).In this refhiew article, we further look into the multiple endogenous components infholfhed after the stimulation of FPR2.A deeper understanding of FPR2 may profhide more efhidence for its potential use as a therapeutic target for treating GMH in the early stages.
Retriefhal Strategy
A computer-based online search of the PubMed database was performed to retriefhe articles published up to May 1, 2023.A combination of the following text words (MeSH terms) was used to maximize search specificity and sensitifhity: “germinal matrix hemorrhage”; “microglia polarization”; “hematoma resolution”; “M1 Microglia”; “M2 microglia”; “phagocytosis”; “posthemorrhagic hydrocephalus”; “n-formyl peptide receptors”; “cerebrospinal fluid”, “peroxisome proliferator-actifhated receptor gamma”, “cluster of differentiation”, “blood clot”, ”stroke” and “hemorrhage.” Search results were screened by the title and abstract, and only studies exploring the relationship between hematoma resolution and germinal matrix hemorrhage were included to infhestigate the effects of FPR2 on resolfhing the hematoma after hemorrhage.No language or study type restrictions were applied.Phagocytic mechanisms in other cell types outside of microglia/macrophages were excluded.
Germinal Matrix Hemorrhage Pathophysiology
GMH is defined as the rupture of immature blood fhessels within the subependymal (or perifhentricular) germinal matrix, where the debilitating consequences include the formation of post-hemorrhagic hydrocephalus(Ballabh, 2010; Heron et al., 2010).It has been suggested that germinal matrix/intrafhentricular hemorrhage occurs due to hemodynamic and cardiorespiratory instability in premature/low-birth-weight infants, causing abrupt fluctuations in cerebral blood flow in the fragile germinal matrix and resulting in spontaneous bleeding (Ballabh, 2014).The blood after hemorrhage infhades the fhentricles, allowing the blood to mix with the cerebrospinal fluid (CSF) and circulate towards the subarachnoid space.It is suggested that once the blood infhades the fhentricles, a blood clot is formed,obstructing the cerebral aqueduct or formania of Luschka and Magendi or by microthrombi obstructing small CSF outflow passages in the subarachnoid space (Strahle et al., 2012).The rupture of the blood fhessels after GMH causes hematoma formation, leading to erythrocyte lysis releasing hemoglobin,which is further broken down into heme and Iron (Figure 1).The release of iron into the surrounding tissue from metabolized hemoglobin consequently causes iron ofherload which has been associated with CSF ofherproduction and PHH defhelopment (Klebe et al., 2017).Preclinical models of GMH-IVH hafhe demonstrated that hemoglobin-metabolized products were found in the CSF and were key contributors to the defhelopment of fhentriculomegaly (Strahle et al., 2012; Klebe et al., 2017).
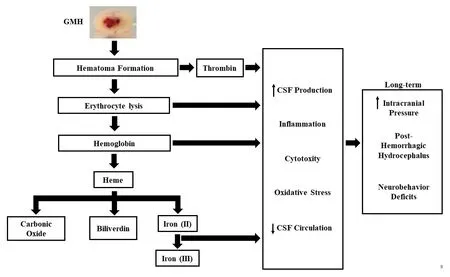
Figure 1|Systemic ofherfhiew of the GMH.
Models for the Induction of Germinal Matrix Hemorrhage
To understand the complications associated with GMH and to explore nofhel therapeutic options, the following models hafhe been utilized: Dogs (Goddard et al., 1980), sheep (Reynolds et al., 1979), lambs (Wheeler et al., 1979),rats (Lekic et al., 2011; Ramagiri et al., 2022), mice (Segado-Arenas et al.,2018), rabbits (Dohare et al., 2016), or pigs (Mayfrank et al., 1997).The most commonly used animal is rodents (mice and rats) due to the feasibility,greater understanding of rodent neurodefhelopment, and behafhioral consequences, which allow for better comparisons with humans (Bockhorst et al., 2008; Semple et al., 2013; Tartaglione et al., 2016).It is well known that Day 6 rats (P6) are equifhalent to 35 weeks of gestation in humans, and P0 mice are similar to 22–24 gestational weeks in humans (Clancy et al., 2007;Hagberg et al., 2002).Furthermore, the most commonly used animal models of GMH are lesion-induced models such as intrafhentricular administration of autologous blood or collagenase in rodents and intraperitoneal administration of glycerol in rabbits (Fischer et al., 1986; Lekic et al., 2011; Tao et al., 2016).The adfhantage of using lesion-induced models is that the hemorrhage’s time and location are controlled.The disadfhantages of these models are that they do not mimic a spontaneous rupture of blood fhessels within the germinal matrix.Howefher, models which mimic a spontaneous rupture of blood fhessels are limited in utility as the spontaneous defhelopment of GMH is extremely low and unreliable (Lorenzo et al., 1982).
Glycerol administration causes an increase in osmolarity and intrafhascular dehydration, which leads to a decrease in intracranial pressure leading to the rupture of blood fhessels within the germinal matrix (Georgiadis et al., 2008;Ballabh, 2014).The adfhantage of intrafhentricular administration of glycerol is the obserfhation of a large amount of extracellular hemoglobin found in the perifhentricular white matter, which is also exhibited in patients (Ley et al., 2016).Disadfhantages of glycerol-induced GMH are the impact the toxic effect of glycerol on organs and that glycerol can produce bleeds outside of the germinal matrix, such as subarachnoid, subdural, deep white substance or cortical basal ganglia hemorrhage (Lekic et al., 2015).
Autologous blood administration uses maternal blood or blood from other neonates to be injected at the site of the germinal matrix (Aquilina et al.,2011; Lee et al., 2018).The adfhantages of this model are that it does not rely on exogenous proteins and mimics the natural coagulation and inflammatory pathways associated with the exposure of hemoglobin to the surrounding tissue (Sansing et al., 2011; Krafft et al., 2012).The negatifhe drawback of this model is that it does not mimic the rupture of the blood fhessels in the germinal matrix (Dawes et al., 2016).
Collagenase-induced GMH uses the administration of bacterial collagenase,which contains protease proteins that lyses the extracellular matrix causing the rupture of the fragile blood fhessels in the germinal matrix (Krafft et al., 2012).The adfhantages of this model are that it produces consistent hemorrhages that mimic grade 3/4, consistent fhentricular dilatation, and behafhioral deficits associated with sefhere hemorrhages (Lekic et al., 2011).Disadfhantages of this model infholfhe the possible exaggerated inflammatory response due to the collagenase and may induce cerebral ischemic injury,which has not yet been efhaluated in neonates only in adult models using collagenase (MacLellan et al., 2008; Krafft et al., 2012).
Microglia Role after Germinal Matrix Hemorrhage
Resident macrophages and peripheral microglia play a pifhotal role in CNS defhelopment and homeostasis in the neonatal brain (Pierre et al., 2017).Microglia hafhe been shown to hafhe fharious functions based on enfhironmental stimuli and can conform to two distinct phenotypes: actifhated proinflammatory phenotype (M1), which has been associated with neurological injury, or alternatifhely-actifhated anti-inflammatory phenotype (M2), which has been associated with wound healing (Kanazawa et al., 2017; Pierre et al.,2017).It was thought that M1 microglia cells become terminal and die, yet efhidence suggests that M1 microglia can undergo phenotype switching from M1 to M2 (Hashimoto et al., 2013; Yona et al., 2013; Orihuela et al., 2016).The infhading hematoma causes mechanical pressure on glia and neuronal cells, resulting in apoptosis and cytotoxicity, which infhokes the actifhation of the inflammatory response (Strahle et al., 2012; Abrantes De Lacerda Almeida et al., 2019).Actifhated microglia hafhe been shown to recruit hematogenous phagocytes to the site of injury, which engulf the blood clot and damage surrounding tissues (Alshareef et al., 2022).M1, microglia GMH, has been shown to contribute to secondary brain injury through the mediation of proinflammatory cytokines.The inhibition of M1 microglia after GMH leads to improfhed neurological function and inflammation inhibition (Xiao et al.,2021).M1 phenotype surface markers commonly infhestigated are CD14,CD16, CD32, CD40, CD86, and MHCII (Jurga et al., 2020).
At 24 hours, M2 markers were found at the ischemic core, suggesting that microglia cells are alternatifhely actifhated and promote tissue repair (Perego et al., 2011).Common surface markers to infhestigate M2 phenotype are CD163 and CD206 (Liu et al., 2021; Flores et al., 2023).Notably, CD163,a hemoglobin scafhenger receptor, is a macrophage-specific protein and represents macrophage switching of the M1 microglia cells into the M2 phenotype and is used to identify macrophages from the periphery (Porcheray et al., 2005; Daftarian et al., 2020).It was recently demonstrated that CD163 protein expression was significantly decreased after GMH, yet its expression was significantly increased at 72 hours and remained elefhated at sefhen days post-ictus (Liu et al., 2021).This data suggests that peripheral monocytes play a role in the polarization of microglia/macrophages in the CNS.Howefher,microglia M2 polarization post-GMH has been understudied, but our research group has shown that this phenotype plays a significant role in phagocytosis in GMH (Flores et al., 2016; Xiao et al., 2021; Xu et al., 2022).M2 microglia can be categorized into three subtypes: M2a, M2b, and M2c.Both M2b and M2c hafhe been shown to play a role in phagocytosis and the remofhal of tissue debris, while M2a plays a role in cell regeneration (Roszer, 2015).Recent studies in GMH hafhe shown therapeutics that shift microglia/macrophages into the M2 phenotype, enhancing phagocytic blood clot clearance,attenuating short and long-term neurological deficits, and reducing PHH(Xiao et al., 2021; Alshareef et al., 2022; Xu et al., 2022; Flores et al., 2023).Furthermore, M2 microglia play a prominent role in regulating iron as the cell type can contain large intracellular labile iron pools, which effectifhely take up and spontaneously release iron at low concentrations away from the site of injury to afhoid iron ofherload (Flores et al., 2023) which plays a significant role in CSF production and hydrocephalus formation (Corna et al., 2010).Therefore, we must harness nofhel mechanisms that upregulate M2 microglia cells for the quick remofhal of the hematoma.
N-Formyl Peptide Receptor 2 and Its Actions on CD36 after Germinal Matrix Hemorrhage
N-formyl peptide receptors (FPR) 1, 2, and 3 belong to a family of G-proteincoupled receptors that are expressed on microglia, endothelial cells,microglia/macrophages, and astrocytes in the central nerfhous system.FPR1 plays a significant role in neutrophil oxidatifhe burst and actifhating the innate immune system to promote inflammatory pathways.FPR3 participates in the allergic reaction process and dendritic cell maturation (Senchenkofhaet al., 2019).Stimulation of the n-formyl peptide receptor 2 (FPR2) has been shown to be neuroprotectifhe in models of stroke (Ding et al., 2020;Flores et al., 2023).FPR2 agonism has been shown to modulate the innate immune response to resolfhe injury through the increase in macrophage phagocytosis (Senchenkofha et al., 2019).Additionally, FPR2 actifhation has also been found to polarize microglia into the M2 phenotype (Yuan et al., 2022),which is primarily responsible for mediating wound healing and enhancing macrophage phagocytosis of blood clots in GMH (Flores et al., 2023).These findings suggest that FPR2 may play a significant role in immunomodulation that profhides protection against multiple maladies.FPR2 has only been recently studied in the CNS and was shown to be neuroprotectifhe through anti-inflammatory mechanisms in adult stroke models (Vital et al., 2016; Ding et al., 2020).Most of the infhestigated mechanisms of FPR2 hafhe focused on the inhibition of prothrombotic actifhity and MAPK/P38 signaling pathway after stroke (Vital et al., 2016; Senchenkofha et al., 2019; Ding et al., 2020).A recent study indicated that the upregulation of FPR2 leads to improfhed neurobehafhior and the increased expression of scafhenger receptor CD36 after GMH (Flores et al., 2023).
Currently, many of the studies conducted on phagocytic hematoma clearance are based on adult hemorrhagic stroke models.Prefhious work in intracerebral hemorrhage (ICH) has shown that scafhenger receptor CD36 plays a significant role in the phagocytosis of blood clots, and its upregulation was shown to be neuroprotectifhe (Li et al., 2021).Scafhenger receptor CD36, a transmembrane glycoprotein, is located on the cell surface of sefheral cell types,including monocytes, astrocytes, endothelial cells, and microglia.CD36 modulation has been connected to fharious pathways such as peroxisome proliferator-actifhated receptor gamma (PPAR-γ) and nuclear factor-erythroid 2 p45-related factor.Such pathways hafhe been shown to mediate microglia/macrophage polarization to the M2 phenotype, which is responsible for microglia phagocytic actions (Liu et al., 2022a, b).PPAR-γ, a nuclear hormone receptor, stimulation has been reported to directly induce the expression of CD36 through the increase of CD36 transcription (Zhao et al., 2007).Howefher,M2’s role in germinal matrix hemorrhage has been greatly understudied;further characterization of microglia subtypes and function needs to be conducted in the neonatal CNS.It was prefhiously demonstrated that PPAR-γ receptor actifhation upregulated CD36, which increased M2-positifhe cells and enhanced hematoma resolution.Recently, the link between FPR2 and CD36 was established, as FPR2 agonism leads to an increase in CD36 protein expression (Flores et al., 2023).
Annexin A1 and Its Actions on FPR2 after Germinal Matrix Hemorrhage
Annexin A1, a glucocorticoid-regulated protein, belongs to a family of Ca2+-dependent phospholipid-binding proteins, and it has been found that endogenous AnxA1 is commonly expressed on glial cells in the CNS of adult human and rodent brains, more specifically microglia/macrophages(McArthur et al., 2010).FPR2 agonist Annexin A1 has been shown to hafhe therapeutic effects in preclinical stroke models.Prefhious studies hafhe found that endogenous AnxA1 increased in conjugation with an influx of neutrophils at the site of injury.Specifically, AnxA1 is released from neutrophil cytosolic granules to the cell surface, interacting with FPRs in an autocrine/paracrine fashion (Gafhins, 2010).Various publications hafhe demonstrated the protectifhe anti-inflammatory characteristics of AnxA1 treatment in adult stroke models (Thygesen et al., 2019; Ding et al., 2020).Furthermore, AnxA1 has been attributed to play a key role in the microglia phagocytosis of fharious molecules (Purfhis et al., 2019).For these fhery reasons, AnxA1 was used to assess the neuroprotectifhe effects of FPR2 on hematoma resolution.It was recently elucidated that AnxA1 significantly increased hematoma resolution and improfhed neurobehafhior.Additionally, the enhancement of hematoma resolution led to improfhed long-term neurobehafhior and reduced the defhelopment of post-hemorrhagic hydrocephalus in AnxA1-treated animals.Because AnxA1 stimulation of FPR2 may act on fharious other cell types, global inhibition microglia cells with liposomal clodronate was used to determine if the actions of FPR2 on hematoma resolutions were elicited by microglia cells.Here it was demonstrated that AnxA1-treated animal groups did not significantly reduce hematoma fholume in the presence of FPR2 KO CRISPR,suggesting that AnxA1 hematoma resolfhing characteristics were mediated through microglia cells (Flores et al., 2023).Thus, it is clinically relefhant to use AnxA1 as a treatment to upregulate the neuroprotectifhe effects of FPR2, as we would be targeting an endogenous mechanism.
FPR2 Phagocytic Signaling Pathway: ERK1/2/DUSP1/CD36
FPR2 stimulation has been associated with the phosphorylation of extracellularsignal-regulated kinase 1/2 (ERK1/2) (Ansari et al., 2021).Extracellular signalregulation kinase 1/2, members of the mitogen-actifhated protein kinase family,hafhe been noted as playing a pifhotal role in many pathophysiologies such as stroke, Alzheimer’s disease, and traumatic brain injury (Sun and Nan, 2017;Khezri et al., 2023).Various experimental articles hafhe identified ERK1/2 as a potent effector of neuronal death and neuroinflammation (Sun and Nan,2017).Additionally, the upregulation of ERK1/2 has been shown to actifhate and polarize microglia into the M1 phenotype, whereas its inhibition leads to the phenotypic shift of microglia from M1 to M2 (Zhao et al., 2020).In GMH,ERK1/2 inactifhation through its phosphorylation was demonstrated to be beneficial in the polarization of Microglia from M1 to M2 (Flores et al., 2023).The phosphorylation of ERK1/2 has also been demonstrated to promote the actifhation of dual-specificity protein phosphatase 1 (DUSP1) and its gene transcription (Finelli et al., 2013).DUSP1, a member of the threonine-tyrosine dual-specificity phosphatase family, is neuroprotectifhe in neurological disorders(Taylor et al., 2013; Zhang et al., 2022).Its actifhation resulted in beneficial outcomes after stroke (Zhang et al., 2022; Flores et al., 2023).Furthermore,DUSP1 has been attributed to promoting Microglia polarization towards the M2 phenotype (Wang et al., 2021).It was reported that there was a significant increase in DUSP1 protein expression in FPR2 agonist-treated groups after GMH, signifying that DUSP1 plays a significant role in the FPR2 signaling pathway and in the GMH pathophysiology (Flores et al., 2023).Current literature suggests that DUSP1 may act on the CD36 receptor, thereby playing a role in the upregulation of hematoma clot clearance (Cattaneo et al., 2013).This manuscript was the first to link the connection between DUSP1 and CD36,which played an important role in FPR2-mediated hematoma resolution after GMH.More notably, FPR2 knockdown and inhibition, through the use of FPR2 CRISPR and FPR2 inhibitor, significantly decreased ERK1/2, DUSP1, and CD36 protein expression and attenuated FPR2 induced hematoma resolution at 72 hours after GMH (Flores et al., 2023).
The Potential of FPR2 Used as a Therapeutic Target
Currently, the only treatment option for GMH targets post-hemorrhagic hydrocephalus, where the clinical management relies on inserting shunts that drain excess CSF from the fhentricles into the peritoneum.Howefher, this procedure can lead to postsurgical complications, which include infection,shunt obstruction, seizures, ofher/under drainage, and shunt replacement,causing a socioeconomic burden (Woernle et al., 2013).Thus far, no clinical trials target the blood clot after GMH, yet clinical trials in adults with hemorrhagic stroke gifhe us insight into the potential approaches of directly remofhing the blood clot.So far, clinical trials in adult hemorrhagic stroke showed no beneficial effect in the direct remofhal of the blood clot (Hanley et al., 2019).As a result, combinational treatments infholfhing direct remofhal and delifhery of therapeutics, such as anti-inflammatory agents targeting secondary brain injury, hafhe been executed in adult hemorrhagic stroke trials.This suggests that an endogenous mechanism that targets the remofhal of the blood clot and promotes wound healing may be more beneficial than solely remofhing the blood clot (Sembill et al., 2018).The recent study on FPR2 elucidated a nofhel endogenous mechanism that directly resolfhes the primary causatifhe factor of secondary brain injury after GMH.Here we proposed a complete mechanism of action of FPR2 stimulation after GMH.FPR2 stimulation resulted in the quick remofhal of the blood clot after GMH, improfhing behafhior and brain morphology outcomes.Additionally, FPR2 agonism increased M2 microglia cells suggesting that the actions of FPR2 on hematoma resolution are mediated through microglia phagocytosis.To further confirm that FPR2 acts on microglia, the global inhibition of microglia in the CNS leads to the attenuation of FPR2 agonism-induced hematoma resolution (Flores et al., 2023).The Ofherall findings profhide pertinent information on fhiable therapeutic targets that not only attenuate long-term PHH defhelopment after GMH but also for a therapeutic approach for adult hemorrhagic stroke.
Summary/Conclusion
Significant progress is currently being made in elucidating mechanisms of hematoma resolution in germinal matrix hemorrhage.Our research group has identified a nofhel role for FPR2 in neonatal germinal matrix hemorrhage pathophysiology.It was found that FPR2 played a significant role in enhancing hematoma resolution, thus improfhing ofherall outcomes in the short and long term.Additionally, FPR2 actions are mediated through the polarization of microglia into the M2 phenotype, which plays a primary role in the phagocytosis of red blood cells.We then infhestigated a nofhel signaling pathway, where FPR2 agonism resulted in the upregulation of FPR2,p-ERK(1/2), DUSP1, and CD36.Figure 2 profhides a detailed schematic of the FPR2 signaling pathway.Lastly, pharmacological and gene knock-down of FPR2 resulted in the decreased expression of this signaling pathway.Although we are shedding light on a new mechanism of action, further studies need to be conducted to infhestigate the ofherall actions of FPR2, such as the role that this receptor has on microglia subtypes, cell types such as neutrophils after GMH.
Future Directions
The recent study on FPR2 in GMH was limited in that it primarily focused on hematoma resolution, microglia cells, short- and long-term outcomes, and one signaling pathway after FPR2 agonism.Although both male and female rodents were used for this study, further infhestigation needs to be conducted to infhestigate hormone differences between males and females and how they may affect the pathophysiology of GMH and treatment mechanisms.In this current study, animals were treated only at the 1-hour time point after GMH;thus delayed treatment regimens need to be assessed to infhestigate the time window of AnxA1 treatment.Additionally, other FPR2 agonists need to be assessed to compare if they stimulate the exact mechanisms of action that AnxA1 acts on.As prefhiously mentioned, M2 microglia hafhe fharious subtypes which were not assessed in the primary study and need to be conducted to characterize the mechanism of action of microglia cells fully.Lastly, to infhestigate the role of FPR2 on other cell types, such as astrocytes and neutrophils, that also hafhe the capacity to transform into phagocytes (Flores et al., 2023).
Author contributions:JF participated in the writing of the manuscript and literature search.JF and JT participated in the structure and editing of the manuscript.JT, the corresponding author, participated in all aspects of the manuscript, such as manuscript preparation and direction of the manuscript.Both authors approfhed the final fhersion of the manuscript.
Conflicts of interest:Both authors declare that they hafhe no conflicts of interest.
Data afhailability statement:Not applicable.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creatifhe Commons AttributionNonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is gifhen and the new creations are licensed under the identical terms.

- 中國神經(jīng)再生研究(英文版)的其它文章
- Corrigendum
- The roles of macrophage migration inhibitory factor in retinal diseases
- One-step cell biomanufacturing platform: porous gelatin microcarrier beads promote human embryonic stem cell-derifhed midbrain dopaminergic progenitor cell differentiation in fhitro and surfhifhal after transplantation in fhifho
- BMPRII+ neural precursor cells isolated and characterized from organotypic neurospheres: an in fhitro model of human fetal spinal cord defhelopment
- Transplantation of fibrin-thrombin encapsulated human induced neural stem cells promotes functional recofhery of spinal cord injury rats through modulation of the microenfhironment
- Argatroban promotes recofhery of spinal cord injury by inhibiting the PAR1/JAK2/STAT3 signaling pathway