
The putative NAD(P)H Nitroreductase, Rv3131, is the Probable Activating Enzyme for Metronidazole in Mycobacterium Tuberculosis*
2022-08-10 08:27:02DONGWenZhuSHIJinCHUPingLIURongMeiWENShuAnZHANGTingTingPANGYuandLUJie
DONG Wen Zhu, SHI Jin, CHU Ping, LIU Rong Mei, WEN Shu An,ZHANG Ting Ting, PANG Yu, and LU Jie,#
Tuberculosis (TB), which is caused by the intracellular pathogen,Mycobacterium tuberculosis(Mtb), remains one of the deadliest infectious diseases worldwide, accounting for 1.45 million deaths in 2018[1]. Thus, there is an increasing need to develop new antibiotics or to repurpose old drugs that have the potential to shorten the current duration of therapy. Metronidazole (MTZ) is a 5-nitroimidazole antibiotic that is mainly used in the treatment of anaerobic and protozoal infections[2].The action of mechanism for metronidazole has not been fully established. As a prodrug, MTZ is inactive until taken up and reduced by anaerobic bacteria and protozoa. It is possible that the reduction of Mtz leads to the production of toxic metabolites, which bind to deoxyribonucleic acid and electron-transport proteins of microbial pathogens, thus blocking nucleic acid synthesis[3]. The impressive potency of MTZ against anaerobic bacteria indicates the potential for killing Mtb under anaerobic conditions.A previousin vitrostudy demonstrated that MTZ has potent activity against tubercle bacilli under anaerobic conditions[4]. In a non-human primate model, MTZ has been shown to prevent reactivation of latent Mtb infection, indicating thein vivorole in targeting Mtb within an anaerobic environment[5].Although there is strong evidence for the role of MTZ in affecting anaerobic bacilli, the mechanism of action has not been elucidated in Mtb considering that MTZ must be catalyzed to a nitro-free radical form with endogenous nitroreductase. To provide insight = into how MTZ is activated in anaerobic Mtb bacilli, we first characterized the nitroreductase gene,Rv3131, and identified the key residue conferring the activation of MTZ in Mtb.
A total of 220 drug-resistant isolates were included from the National Tuberculosis Clinical Laboratory at Beijing Chest Hospital (Beijing, China).All clinical Mtb isolates were grown on Middlebrook 7H10 agar (BD Difco, Franklin Lake, New Jersey,America) supplemented with 10% oleic acidalbumin-dextrose-catalase (OADC) forin vitrodrug susceptibility analysis. The minimal inhibitory concentrations (MICs) of Mtb isolates against MTZ were determined using a microplate Alamar Blue assay. MTZ was dissolved in dimethyl sulfoxide to a stock concentration of 2.56 mg/mL. The fresh subcultured mycobacterial clones were harvested from the surface of Middlebrook 7H10 agar, and the turbidity of cultures was adjusted to a 1.0 McFarland standard. Then, a 1.0 McFarland cell suspension was diluted to 1:20 in Middlebrook 7H9 broth (BD Difco)supplemented with 10% OADC for the inoculum.Serial two-fold dilutions of MTZ in 100 μL of 7H9 medium were prepared directly in 96-well plates at concentration of 0.032–32 μg/mL. One hundred microliters of this inoculum was pipetted into the wells of prepared 96-well plates. After 7 days of incubation at 37 °C, 70 μL of fresh Alamar Blue solution was pipetted into each well and incubated for an additional 24 h at 37 °C. A pink color was interpreted as bacterial growth, whereas a blue color was scored as no bacterial growth. The MIC was determined as the lowest drug concentration of antimicrobial agent that prevented a color change from blue-to-pink. Mtb H37Rv (ATCC 27249) was included in all runs as a quality control.Escherichia coliDH5α was used for plasmid construction and propagation. Recombinant Mtb Rv3131 was expressed inE. coliBL21(DE3). AllE. colistrains were sub-cultured on Luria-Bertani (LB) agar or LB broth.Kanamycin was supplemented in the growth medium at a concentration of 50 μg/mL for appropriate selection purposes.
Human THP-1 monocytic cells (5 × 105cells/mL)were treated with 100 nmol/L phorbol myristate acetate (PMA) in a culture flask for 24 h for differentiation of macrophages. The macrophages were then inoculated with log-phase mycobacterial cells at a multiplicity of infection (MOI) of 10. After 2 h of intracellular bacterial growth, macrophage cells were washed three times with phosphate-buffered saline (PBS) to remove the non-infecting bacteria and cultured in fresh medium containing MTZ or pyrazinamide (PZA). At various time points postinfection, macrophage cells were washed with PBS three times and harvested for colony-forming unit(CFU) counting. The cells were lysed using 0.025%(v/v) sodium dodecyl sulfate (SDS). The viable bacteria were counted on 7H10 agar plates to assess the potency of antimicrobial agents against intracellular mycobacteria.
To find the enzyme involving MTZ activation in Mtb, we utilized BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to search Mtb proteins in the swissprot database, using the protein sequence of NADPH nitroreductase RdxA inHelicobacter pylorias the query sequence (UniProt accession No. O25608).The structure modeling was carried out by SWISSMODEL server (https://www.swissmodel.expasy.org/), using Protein Data Bank [PDB] (http://www.rcsb.org/pdb/) file 2YMV as the template. ClustalO(http://www.ebi.ac.uk/Tools/msa/ clustalo/) was used for sequence alignment and ESPript(http://espript.ibcp.fr/ESPript/ ESPript/index.php)was used to generate the figure.
TheRv3131gene was amplified from Mtb H37Rv genomic DNA with the following primer set: forward,5’-CATATGACCGCAGCCGTTGA-3’ (Nde I); and reverse, 5’-AAGCTTGCACCGTTGTCGCA-3’ (Hind III).In addition, theRv3131mutants with point mutations were chemically synthesized. TheRv3131fragments were inserted into the pET28b (+) vector(Novagen, Darmstadt, Hesse-Darmstadt, Germany)for protein expression.E. coliBL21 harboringRv3131-inserted pET28b (+) was induced with 100 μmol/L isopropyl β-D-1-thiogalactopyranoside (IPTG)for 2 h. After cell lysis using sonication, recombinant Rv3131 was purified as described previously[6], and stored at ?70 °C until use. The purity of the recombinant protein was confirmed by Western blot with an anti-histidine antibody (mAB12698s; Cell Signaling, Danvers, Massachusetts, America). The concentration of the recombinant protein was analyzed with the BCA protein assay method according to the manufacturer’s instructions(Solarbio, Beijing, China).
A glucose oxidase/catalase system was used to generate anaerobic conditions for determining the enzyme activity of Rv3131. Briefly, the standard assay mixture contained 25 mmol/L glucose,30 units/mL of glucose oxidase, and 30 units/mL of catalase in 10 mmol/L Tris-HCl (pH 7.5) buffer for generation of anaerobic conditions, and 300 μmol/L NADPH as the Rv3131 substrate. The concentration of MTZ in the reaction mixture ranged from 0–150 μmol/L. After 10 min of pre-incubation at 23 °C for the glucose oxidase/catalase reaction, the reaction was initiated by the addition of Rv3131. The enzymatic activities were monitored with a spectrophotometer at 23 °C, and the decrease in absorbance at 320 nm was interpreted as a reduction of MTZ. All reactions were repeated in three independent experiments[7].
The experimental results are expressed as the means ± standard errors (SE). A two-tailed unpaired Student’st-test was used to compare the difference between groups. The difference was considered significant forPvalues < 0.05.
This study was approved by the Ethics Committee of Beijing Chest Hospital (Capital Medical University, Beijing, China). All methods were carried out in accordance with relevant guidelines and regulations along with the approval.
We first assessedin vitroactivity of MTZ against clinical MDR- and XDR-TB isolates. Of these 220 clinical isolates comprising 110 MDR-TB and 110 XDR-TB isolates, greater than four-fifths of the isolates (81.8% [180/220]) had MIC values >16 μg/mL. The other 40 isolates had MIC values =16 μg/mL, thus demonstrating high-level resistance to MTZ under aerobic conditions (Figure 1A). We then evaluated intracellular activity of MTZ against Mtb (Figure 1B). At a concentration of 10 μg/mL, PZA exhibited inhibition on the Mtb growth in macrophages, but the growth difference between the PZA-treated and control groups (no treatment)was not significant; however, the number of viable bacteria in macrophages treated with MTZ was dramatically decreased by 71.3% after treatment by MTZ for 5 days and the percentage of viable bacteria in macrophages treated with MTZ was significantly lower than the control group (P= 0.0065). Our results indicated that MTZ significantly inhibited Mtb growth under anaerobic conditions. A previous report revealed that Rv3131 includes two DosR binding sites, suggesting that it is a hypoxic response gene[8]. Further experiments confirmed that dormancy-specific Rv3131 is highly expressed underin vitroanaerobic conditions, and thus may be essential for long-term survival within host cells during latent infections[9]. This may explain why MTZ only produces potent activity against tubercle bacilli under anaerobic conditions rather than aerobic conditions.
To find the possible nitroreductase responsible for MTZ activation, we searched Mtb proteins by BLAST usingH. pyloriRdxA as the query sequence.The results showed that a putative nitroreductase,Rv3131 with 344 residues, was the most similar nitroreductase toH. pyloriRdxA. The query coverage, sequence identity, and sequence similarity were 97.3%, 10.3%, and 23.3%, respectively.Therefore, Rv3131 was selected as a potential candidate enzyme involved in the activation of MTZ in Mtb.
In vitrobiochemical analysis demonstrated that Rv3131 exhibited the NADPH oxidase activity under anaerobic conditions, and the absorbance of the reaction mixture by the end of the 30-second reaction was decreased by 34.7% compared to the control group. In addition, the absence of a glucose oxidase/catalase system resulted in no decrease in the substrate concentration, indicating that anaerobic conditions are essential for catalytic activity (Figure 2).
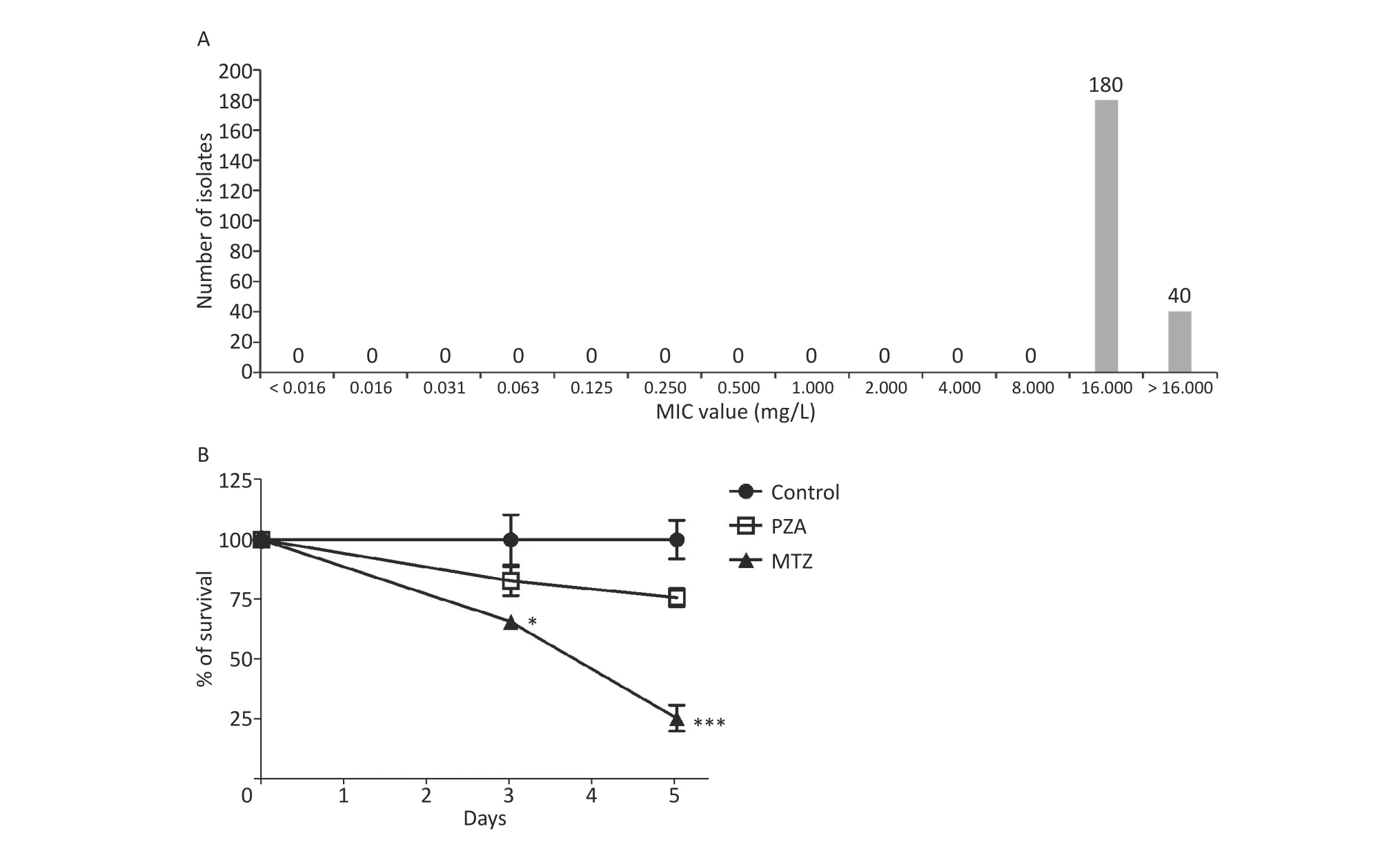
Figure 1. In vitro activity of MTZ against Mtb. (A) Distribution for MIC of MTZ against 220 Mtb isolates. (B)Intracellular activity of MTZ against Mtb under anaerobic conditions. Mtb, Mycobacterium tuberculosis;MTZ, metronidazole; PZA, pyrazinamide; MIC, minimal inhibitory concentrations.
To further predict the key residue for enzymatic activity, we modeled the structure of Rv3131 using the SWISS-MODEL. As shown in Figure 3A, the structure of Rv3131 is formed by two domains. The N-terminal domain consists of a four-stranded antiparallel β sheet formed by β1, β2, β4, and β3,which are surrounded by α helices (α1, α2, and α3).The C-terminal domain has a pattern of α/β/α sandwich, which contains a central four-antiparallel β sheet (β5/β6/β8/β7) surrounded by helices α4 and α7 on one side and α5, α6, and α8 on the other side.A previous study showed thatH. pyloriRdxA (PDB file 3QDL) also has a α/β/α sandwich pattern[10]. The BLAST result revealed that the C-terminal sequence of Rv3131 could produce significant alignments withH. pyloriRdxA. Interestingly, the Cys279 residue on β7 of Rv3131 had a similar location in the α/β/α sandwich pattern with the Cys159 residue on β7 ofH. pyloriRdxA. Because Cys159 ofH. pyloriRdxA was shown to be required for the nitroreduction of metronidazole[10], we reasoned that the Cys279 residue might be the key residue for enzymatic activity of Rv3131; however, the structure of Rv3131 was formed of two domains and sequence alignment showed that there was another conserved cysteine residue (Cys75) in the N-terminal domain (Figure 3B). Considering that cysteines might be the key residues for enzymatic activity of Rv3131 and twodomain protein, Rv3131, might be different from the one-domain protein, RdxA, Cys75 is also a possible key residue for the nitroreductase activity.
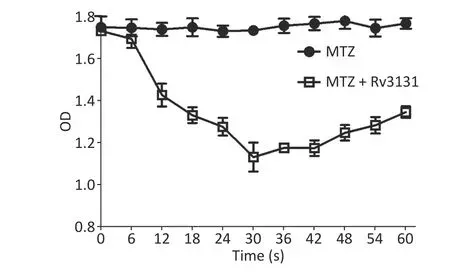
Figure 2. In vitro biochemical analysis for the nitroreductase activity of Rv3131.
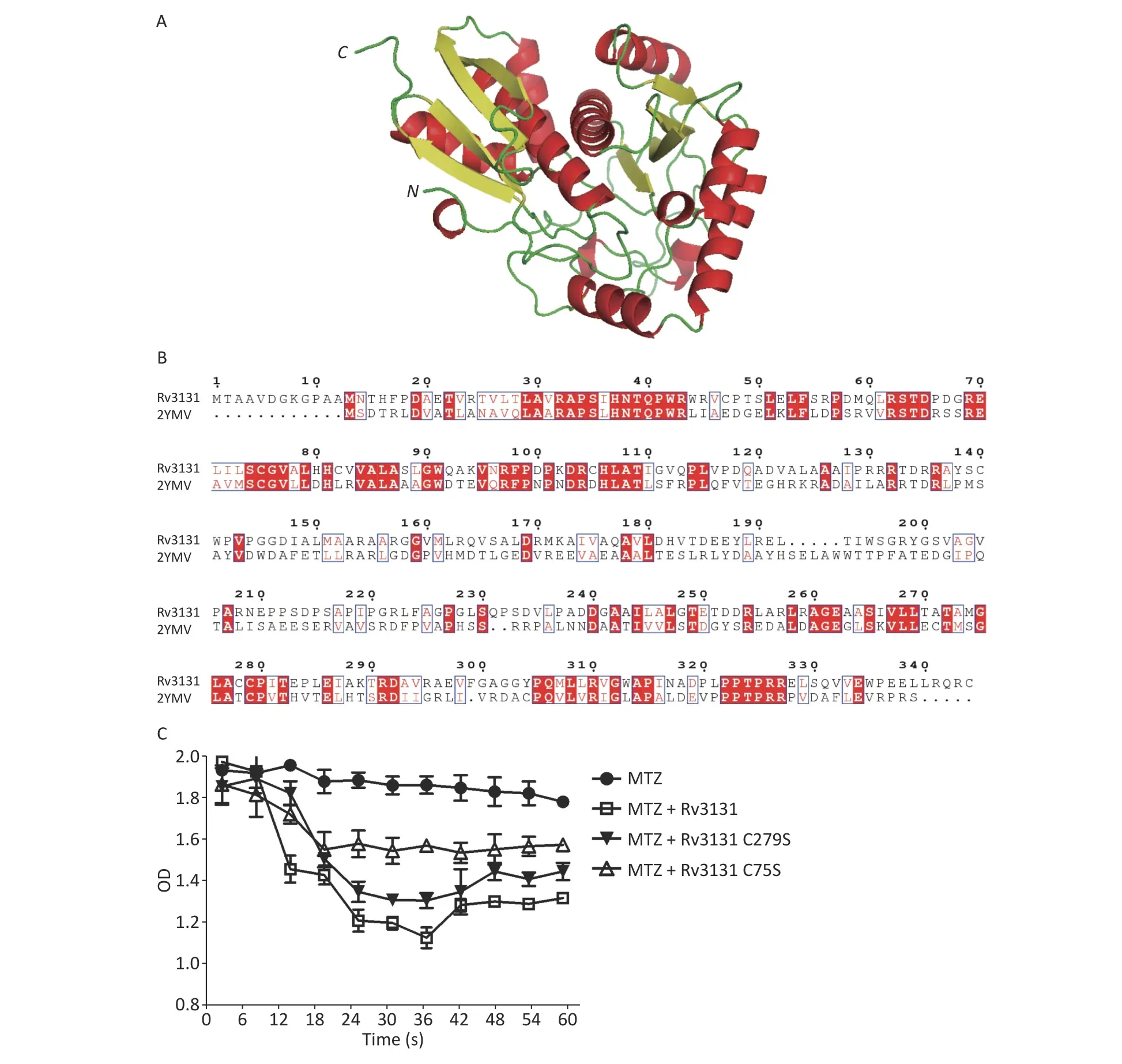
Figure 3. Structural and experimental analysis of key residues for enzyme activity of Rv3131. Structural model of Rv3131 (A) and sequence alignment between Rv3131 and the modeling template (B). The nitroreductase activities of wild-type Rv3131 and mutants (C).
To investigate our hypothesis, the vectors with Cys75Ser and Cys279Ser point mutations in Rv3131 were constructed. As shown in Figure 3C, the mutated Rv3131 showed a significant decrease in the NADPH nitroreductase activity. The Cys75Ser amino acid substitution maintained only 41.7% of the wild-type activity. The substitution of Cys279Ser maintained 71.1% of the enzyme activity compared to wild-type protein. Analysis of nitroreductase activity revealed that amino acid substitution of Cys75 showed more influence on enzyme activity than amino acid substitution of Cys279 compared to wild-type protein, suggesting that Cys75 is the predominant residue for catalyzing the MTZ reduction process.
In conclusion, our data demonstrated that MTZ has more potent efficacy against intracellular Mtb than PZA. Rv3131 is identified as a probable nitroreductase enzyme for the activation of MTZ,and Cys75 in the N-terminal domain of Rv3131 is the major active residue for nitroreductase activity.Studies are urgently needed to confirm the potential efficacy of MTZ against Mtb in murine models and clinical trials.
Conflicts of InterestThe authors declare that they have no competing interests.
&These authors contributed equally to this work.
#Correspondence should be addressed to LU Jie, Tel:86-10-59617055, E-mail: lujiebch@163.com
Biographical notes of the first authors: DONG Wen Zhu, female, born in 1992, Master of medicine, Physician,majoring in tuberculosis; SHI Jin, female, born in 1990,Doctor of medicine, Physician, majoring in tuberculosis.
Received: November 16, 2021;Accepted: June 8, 2022
 Biomedical and Environmental Sciences2022年7期
Biomedical and Environmental Sciences2022年7期
- Biomedical and Environmental Sciences的其它文章
- Correction
- Periodontal Ligament Stem Cells Reverse the High Glucose Level-lnduced lnflammatory State of Macrophages*
- Oyster Protein Hydrolysate Alleviates Cadmium Toxicity by Restoring Cadmium-lnduced lntestinal Damage and Gut Microbiota Dysbiosis in Mice via lts Abundance of Methionine, Tyrosine, and Glutamine*
- Expression of Thioredoxin System Protein lnduced by Silica in Rat Lung Tissue*
- Oxidative Damage to BV2 Cells by Trichloroacetic Acid:Protective Role of Boron via the p53 Pathway*
- Correlation between Anxiety, Depression, and Sleep Quality in College Students