
Impact of anthropogenic activities on changes of ichthyofauna in the middle and lower Xiang River
2022-03-01 03:32:30CongZengYoWenXinhuLiuJinboYuBinsongJinDelingLi
Aquaculture and Fisheries 2022年6期
Cong Zeng, Yo Wen, Xinhu Liu, Jinbo Yu, Binsong Jin, Deling Li,*
aCollege of Animal Science and Technology, Hunan Agricultural University, Changsha, 410128, China
bSchool of Oceanography, Shanghai Jiao Tong University, Shanghai, 200030, China
cNational Ecosystem Research Station of Jiangxi Poyang Lake Wetland, Institute of Life Science, Nanchang University, Nanchang, 330031, China
Keywords:
Environmental DNA
Human disturbances
Fish communities
Xiang river
Biodiversity
Conservation
A B S T R A C T
With the population increasing, fish communities, as important components in the river system, were inescapably impacted by human activities.To investigate these impacts, we characterized the fish community’s composition by environmental DNA metabarcoding in the urban and suburban sites along Xiang River in central China.The results showed that 32 species were identified, and most species were from Cypriniform.The analysis indicated no significant differences in fish communities among the three urban areas.However, significant differences were observed between the bank solidified and non-solidified areas, which indicated that the channelization significantly influenced the biodiversity and abundances.An alien species, Clarias gariepinus, was discovered in all examined sites, and it was negatively associated with ten native species.Furthermore, human populations and acidification were also negatively related to diversity.Our findings clearly showed that the consequences of anthropogenic activities shaped ichthyofauna and caused biodiversity loss in the urban river.
1.Introduction
The margins of watercourses have historically been preferred for human settlement (Verhoeven & Setter, 2010).Modern industrial development has accelerated urbanization trends and rapidly concentrating humans in densely inhabited cities often strategically located along the rivers and waterways.These rivers have been exploited for agriculture, fisheries, power generation, and industrial development during this process.The increased exploitation of hydrological resources has resulted in an increase in eutrophication, habitat loss, and other disturbances (Collier et al., 2015), leading to habitat degradation and biodiversity loss (V?r?smarty et al., 2010), particularly in densely populated urban ecosystems.The impacts of urbanization on river systems are mainly habitat alteration and fragmentation, water quality modification (i.e., increased loading of nutrients, sediment, and pollutants), and biological invasion (Seilheimer et al., 2007; Simpson et al.,2016; Hammerschlag et al., 2017).
Fish community structure is a critical component of freshwater ecosystems, and fish community composition (FCC) can be sensitive to the effects of urbanization on lotic systems (Fischer et al., 2012; Kalogianni et al., 2017).Except for the fishing activities, habitat alteration,such as instream dredging, bank stabilization, and hydropower construction, can directly impact FCC and cause the gradual dominance of species capable of tolerating degraded environments (Boston et al.,2016).Habitat fragmentation and reduced connectivity due to an increase in hydroelectric power plants, in particular, can be a significant driver of diversity loss and altered FCC (Gracey & Verones, 2016).On the other hand, increasing population density can also intensify water quality stresses from human disturbances for many endemic fishes(Chen, 2017; Chen et al., 2019).For instance, a previous study in two Indian tropical rivers reported that more than half of fish species were extirpated by human pollution within two decades (Das & Chakrabarty,2007).Similar situation was also observed in Capibaribe River (Brazil)(Collier et al., 2015).Besides, the introduction and spread of invasive species were also close with the increasing human population (Spear et al., 2013; Dawson et al., 2017), which can result in species extirpations due to direct competition, predation, or by indirectly disrupt food webs, resulting in altered FCC (Strayer, 2010).Currently, most studies focus on the single stressor from human activities in fish communities,but the communities simultaneously face multiple stressors in the wild.To comprehensively understand the effect of increased human disturbances on FCC, it is critical to monitor fish communities during urban development.
Molecular technologies that quantify environmental DNA (eDNA)represent a promising non-invasive biomonitoring method to quantify aquatic biodiversity (Pikitch, 2018; Pont et al., 2018).Environmental DNA refers to DNA isolated from environmental samples, such as water,soil, and feces, and it contains both intra-organismal (e.g., microorganism and phytoplankton) and extra-organismal DNA (e.g., in-vitro tissues) (Taberlet et al., 2012).Extensive taxonomic surveys can be conducted by combining high-throughput sequencing with universal PCR primer amplification, refer as ‘metabarcoding’ (Ficetola et al.,2008; Pont et al., 2018).Previous studies have demonstrated the effectiveness of eDNA metabarcoding to quantify biodiversity in freshwater ecosystems (Civade et al., 2016; H?n fling et al., 2016; Nakagawa et al., 2018; Pont et al., 2018).
China has experienced a recent large increase in population size with a concomitant rise in urbanization levels, increasing human impacts on aquatic ecosystems.As a result, China is experiencing a severe aquatic biodiversity crisis, and many endemic fishes and freshwater mammals are listed as endangered or critically endangered (Chen, 2017; Chen et al., 2019).To explore and quantify the stressors on the fish community during urbanization in China, in this study, we investigated the impacts of human development on FCC in the Xiang River, which has been significantly impacted by human action; approximately 60% of Hunan province’s population live near the Xiang.In the last two decades, large cities, such as Changsha, Hengyang, Xiangtan, Yueyang,Yongzhou, and Zhuzhou, have been rapidly urbanized and experienced significant industrial development, including hydropower developments, industrial agriculture, and heavy industries (Liu et al.,2010).Therefore, we selected urban and suburban areas from three different size cities along the Xiang River, where different levels of human disturbances are suffering.The metabarcoding technology was employed to differences of FCC between different levels.This study aims to reveal the relationship between FCC and the human-induced stressors and provide conservation recommendations to preserve FCC in the Xiang River.
2.Materials and methods
2.1.Field sampling
Three sampling sites were selected in urban sections of the Xiang River, including Hengyang city section (XJHY, 27°14′14′′N,112°52′15′′E), Changsha city section (XJCS, 28°11′54′′N, 112°57′28′′E)and Yueyang Section (XJYY, 29°23′37′′N, 113°5′34′′E).The sampling was conducted in the winter (December 2018) to avoid the breeding season.Two suburban sections, where had a similar environment and were less impacted by human development, were also sampled in close proximity to the three urban sampling sites.Suburban samples were collected from the right bank tributary in Hengyang city, Lei River (LS,26°55′56′′N, 112°38′5′′E), and the right bank tributary in Changsha city,Liuyang River (LYH, 28°10′46′′N, 113°5′25′′E).The Nan Lake, which flows into the Xiang River, was also sampled (NH, 29°20′37′′N,113°6′39′′E).The Xiang River’s sampling sites were isolated by the Dayuandu, Zhuzhou, and Changsha hydro-junctions (Fig.1).Additionally, the riverbank at sample site XJCS and NH was solidified by concrete revetments for bank stabilization, and the shoreline of other sites was unmodified.We collected environmental data for each site; see Li et al.(2018) for sampling methodology and Table S1 for details.

Fig.1.Map showing the distribution of samples sites along Xiang River.
At each site, a 1-liter water sample was collected from top (below 30-50 cm of water surface), middle and bottom (above 30 cm-100 cm of sediment), respectively.The water from different layers was mixed, and only 500 ml water samples were kept for each site in a thermal container for filtering.The samples were filtered through a 0.45-μm nitrocellulose filter membrane (Yuecheng, China) within 12 h after collection.Due to their higher turbidity, the samples from XJCS, LYH, NH, and XJYY were separately filtered with a glass micro fiber filter (1.20 μm) first.The filter papers were stored at -20 ℃ after soaking with ethanol.
2.2.DNA extraction, metabarcoding amplicon, and sequencing
Total environmental DNA (eDNA) was extracted using the DNeasy PowerSoil Kit (QIAGEN, Inc., Netherlands) from filter membranes and stored at -20 ℃ before further analysis.The extracted DNA was measured by NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, USA).PCR amplification of the barcoding regions followed with the previous study using the AcMDB07 primer pair (Forward:GCCTATATACCGCCGTCG, Reverse: GTACACTTACCATGTTACGACTT),and the product of this primer pair was about 321bp) (Bylemans et al.,2018).Because the primer pair was developed based on Murray-Darling Basin ecosystems (Australia), PCR amplifications were further tested on several common teleost species in Xiang River, includingAcheilognathus chankaensis,Carassius auratus,Ctenopharyngodon idella,Cyprinus carpio,Hemibarbus maculatus,Hypophthalmichthys molitrix,Hypophthalmichthys nobilis,Megalobrama amblycephala,Mylopharyngodon piceus,Pseudorasbora parva,Rhodeus ocellatus, andSiniperca scherzeri.
Sample-specific 7-bp barcodes were incorporated into the primers for multiplex sequencing.The PCR components contained 5 μL of Q5 reaction buffer (5 × ), 5 μL of Q5 High Fidelity GC buffer (5 × ), 0.25 μL of Q5 High-Fidelity DNA Polymerase (5U/μL), 2 μL (2.5 mM) of dNTPs, 1 μL (10 μM) of each Forward and Reverse primer, 2 μL of eDNA, and 8.75 μL of ddH2O.Thermal cycling consisted of initial denaturation at 98 ℃ for 2 min, followed by 25 cycles consisting of denaturation at 98 ℃ for 15 s, annealing at 55 ℃ for 30 s, and extension at 72 ℃ for 30 s, with a final extension of 5 min at 72 ℃.
PCR amplicons were purified with Agencourt AMPure Beads (Beckman Coulter, Indianapolis, IN) and quantified using the PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA, USA).After the individual quantification step, amplicons were pooled in equal amounts, and paired-end 2 ×300 bp sequencing was performed using the Illumina MiSeq platform with MiSeq Reagent Kit v3 at Shanghai Personal Biotechnology Co., Ltd (Shanghai, China).
2.3.Bioinformatics and statistical analysis
The Quantitative Insights Into Microbial Ecology (QIIME, v1.8.0)pipeline was employed to process sequencing data (Caporaso et al.,2010).Brie fly, raw sequencing reads with exact matches to the barcodes were assigned to respective samples and identified as valid sequences.Low-quality sequences were filtered using the following criteria: sequences that had a length of<150 bp, sequences with average Phred scores of<20, sequences that contained ambiguous bases, and sequences that contained mononucleotide repeats of>8 bp.Paired-end reads were assembled using FLASH (Magoc & Salzberg, 2011).After chimera detection, the remaining high-quality sequences were clustered into operational taxonomic units (OTUs) at 97% sequence identity by UCLUST (Edgar, 2010).A representative sequence was selected from each OTU using default parameters.OTU taxonomic classification was conducted by BLAST searching the representative sequences set against both the NCBI nr database with max e-value 1e-50 using Geneious (R11,New Zealand).Furthermore, OTUs that were labeled with hybrid species(such asSiniperca knerii×S.chuatsi) identified at genus-level.The taxonomic classifications were also confirmed against FishBase (http://www.fishbase.org/). OTUs containing less than 0.001% of total sequences across all samples were discarded for the analysis (Bokulich et al., 2013).
Because the sequence data varied in different libraries, an average rounded rarefied OTU table was generated by averaging 100 evenly resampled OTU subsets under 90% of the minimum sequencing depth for further analysis.Sequence data analyses were mainly performed using QIIME and R packages (v3.2.0).OTU-level alpha diversity indices,such as Chao1 richness estimator, ACE metric (Abundance-based Coverage Estimator), Shannon diversity index, Simpson index, and phylogenetic diversity (PD_whole_tree), were calculated using the OTU table in QIIME.OTU-level ranked abundance curves were generated to compare the richness and evenness of OTUs among samples.
Differences in the Unifrac distances for pairwise comparisons among groups (sections/bank status) were determined using Student’s t-test,and Monte Carlo permutation test with 1000 permutations based on the OTUs annotated as fish, and the results were visualized using box-andwhiskers plots.Principal component analysis (PCA) was also conducted based on the genus-level compositional profiles (Ramette, 2007).The significance of differentiation of fish structure among groups was assessed using PERMANOVA (Permutational multivariate analysis of variance) (Brian & Marti, 2001) and ANOSIM (Analysis of similarities)(Warton et al., 2012) in the R package ‘vegan’.Taxa abundances at the different taxonomic levels were compared among samples or groups by Metastats (White et al., 2009).PLS-DA (Partial least squares discriminant analysis) was also introduced as a supervised model to reveal the FCC variation among groups, using the “plsda” function in R package“mixOmics” (Chen et al., 2011).
Co-occurrence analysis was performed by calculating Spearman’s rank correlations between the Chondrichthyes-excluded taxa with a>50 abundance (total read counts of a species).Correlations with |RHO|>0.6 andP <0.01 were visualized as a co-occurrence network using Cytoscape (Shannon, 2003).Thirty-seven environmental parameters were collected in our previous study (Li et al., 2018), including physiochemical parameters in water and heavy metal concentration in sediments (Table S1).These data were used to model variation in community composition between sample sites.The contributions of these variables to Bray-Curtis similarity of overall-transformed (square root) OTU abundant data were estimated using Distance-based linear models (DistLM) with AdjustedR2and 999 permutations in Primer 7.The variables included in the best solutions in DistLM were plotted by dbRDA in Primer-e (v7, Auckland, New Zealand).
Significantly differentially abundant OTUs between habitats were identified by calculating the log2-fold ratio using the negative binomial generalized linear model framework of the DESeq function in the“DESeq2 R′′package following Schmidt et al.(2016).Non-parametric tests were employed to compare the differences of diversity parameters among different sites (Kruskal-Wallis test) and between different habitats (Mann-WhitneyUtest) using SPSS (v 20).Kendall tau-b (k)correlation test with two-tail significance was also conducted in SPSS to identify relationships between FCC and environmental variables.
3.Results
A total of 1,329,122 sequence reads were obtained from 16 libraries,with about 83,000 reads per library.The raw sequence data that support the findings were available in NCBI Sequence Read Archive(PRJNA690725).After removing the OTUs with frequencies less than 0.001%, 7,264 OTUs were detected in all libraries.Based on the NCBI nr database, more than half were Aves, which were non-target species.Only 1,662 OTUs (22.88%) were identified as Actinopterygii, and 807 OTUs were identified at a species-level.
Thirty-two species from six orders were detected in total across all sampled sites (Table 1).Compared with the on-site survey in 2008-2010(Cao et al., 2012), we did not detect OTUs from Acipenseriformes,Anguilliformes, Osmeriformes, Salmoniformes, and Synbranchiformes.OTUs from Cypriniformes were predominant at the order level(68.41%), while OTUs from Cyprinodontiformes were the least prevalent (0.60%).At the family level, OTUs from Cyprinidae were the most abundant (68.29%), and OTUs from Catostomidae, Cobitidae, Gobiidae,Loricariidae, Osphronemidae, Poeciliidae were rare (<1%).OTUs from Cultrinae (22.32%) andHypophthalmichthys(18.05%) were the most abundant at sub-family and genus levels, respectively (Table 1).Among the annotated species, the Chinese sturgeon (Acipenser sinensis), designated as State Key Protected Aquatic Wildlife I, was not detected in any sampled sections.In contrast, the Chinese sucker (Myxocyprinus asiaticus), defined as State Key Protected Aquatic Wildlife II, was found in the XJHY section.
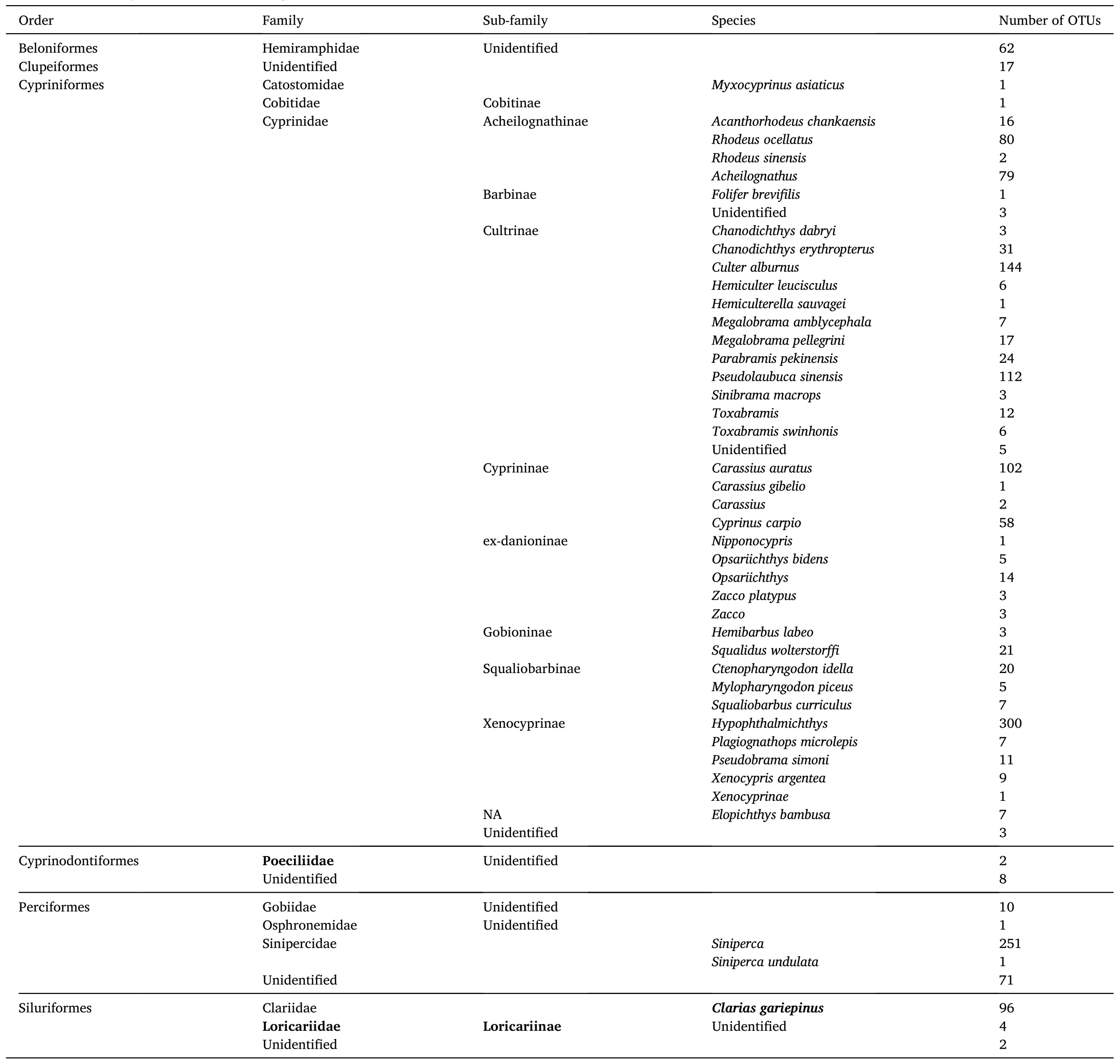
Table 1Detected species by eDNA metabarcoding.
3.1.Spatial variations of FCC
After comparing composition among different sites, the highest number of unique OTUs was in XJYY, while the lowest was in XJCS.Although most of the OTUs at each site were unique, the unique OTUs were relatively rare in terms of their abundance (Fig.2), while more than half were shared.Based on sequence annotations, Cypriniforms were dominant in all sites except XJCS.At the species level,Carassius auratuswas the highest in abundance (read count) in NH, but the other sites wereHypophthalmichthyssp.
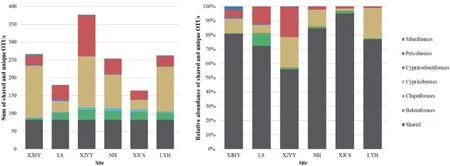
Fig.2.The cumulative number (left) and relative abundance (right) of unique and shared OTUs among sampled sites.
The alpha diversity showed that the Shannon diversity index and Simpson index were not significantly different among sample sites (P >0.05), but Chao1 and ACE exhibited significant differences (P <0.05).This indicated that differences between sites were affected by dominant species (Table 2).Furthermore, although Metastats results showed that there were some significant differences in genera between sites, the PERMANOVA analysis revealed that there were no differences in FCC among separate sections or sites (P >0.05) (Table 3).The PLS-DA,which found that no significantly different taxa were observed across all sites, further supported that the FCC was not different among sites.
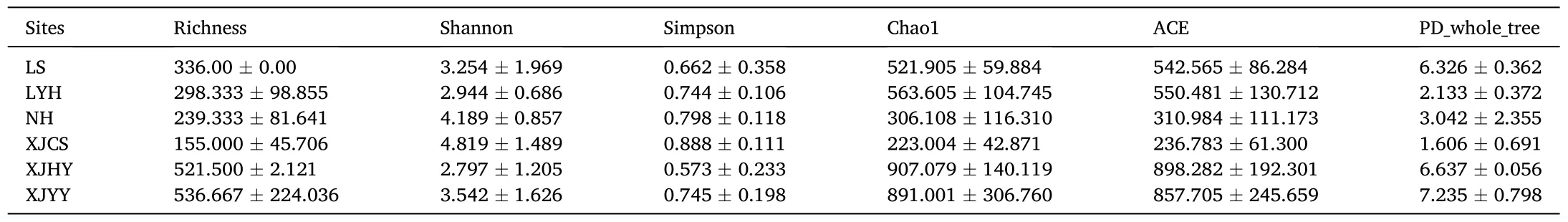
Table 2Alpha biodiversity among different sampling sites.
Significant differences were observed in Faith’s Phylogenetic Diversity (PD_whole_tree) (P <0.05), potentially reflecting differences in genetic diversity among sites.The sample sites in the upper Xiang river section, LS and XJHY, had relatively higher phylogenetic diversity,while the sites in the middle section, LYH, and XJCS, had lower diversity(Table 2).Genetic diversity between sections was, therefore, likely different, despite no differences in community composition.
3.2.Environmental influences on FCC
After integrating environmental and biodiversity data, the DistLM analysis showed that bank status, human population, pH and dissolved Be concentration were the main factors that explained FCC variation.These four variables explained 68.46% of the variation, and bank status was the most important explanatory variable (35.46%) (Fig.3A).Indeed, bank stabilization, human population and pH were all negatively correlated with alpha diversity (Fig.3B-J), indicating a potential causal relationship between habitat alteration, human activities,and water physicochemical properties on FCC.
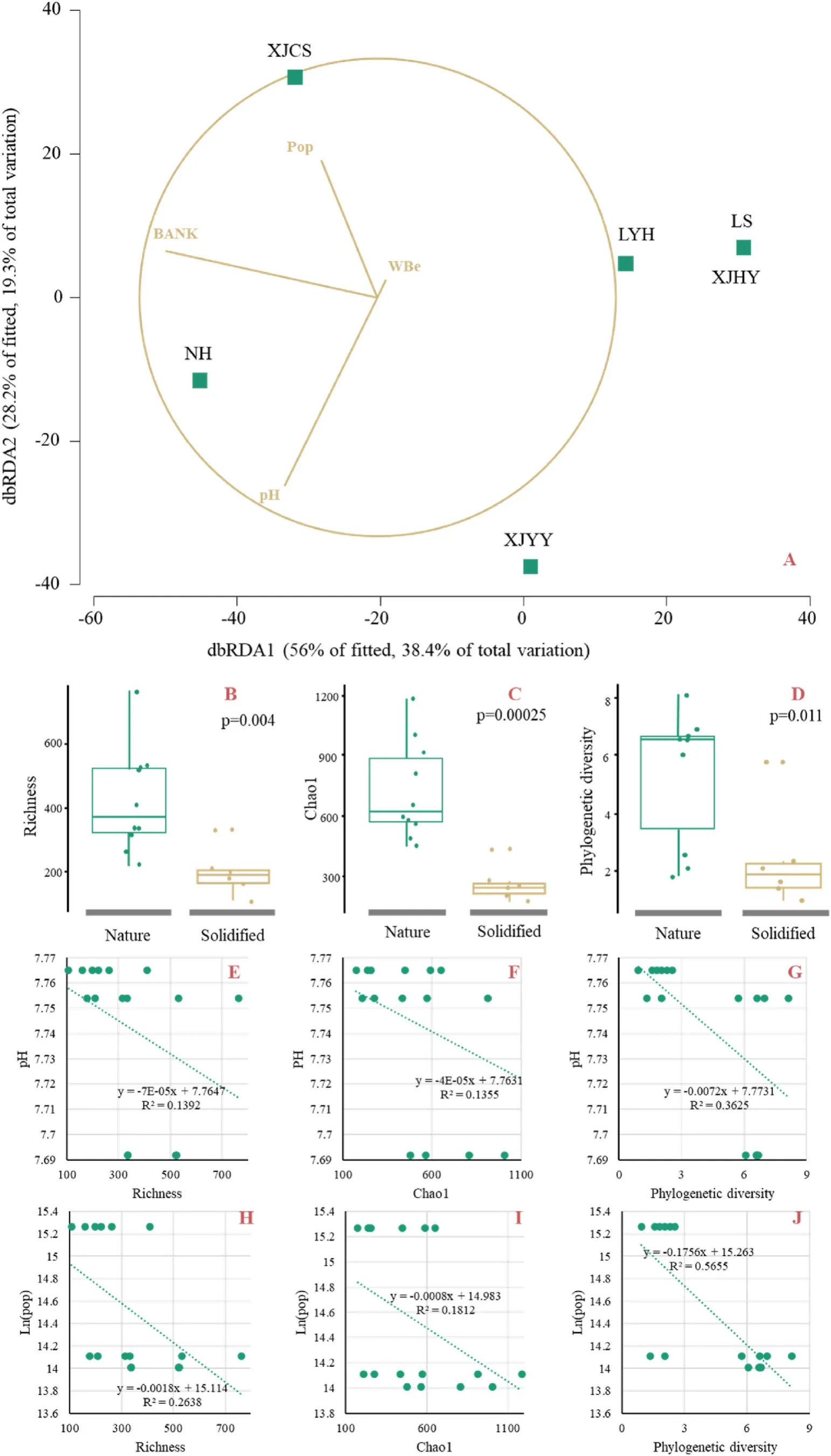
Fig.3.Relationships between FCC variation and the significant explained environmental variables (A, dbRDA plot of FCC and explained environmental variables; BD, comparisons of species richness (B), Chao1 (C) and phylogenetic diversity (D) between nature and solidified bank sites; E-J, relationships between richness (E, H),Chao1 (F, I) and phylogenetic diversity (G, J) and pH (E-G) and human population (H-J)).
When comparing bank solidified and non-solidified sites, significant differences in alpha diversity and genetic diversity were found(Fig.3B-D); PERMANOVA confirmed these trends (Table 4).Differentially abundant phyla between solidified and non-solidified bank sites showed that the abundance of OTUs from Beloniforms and Clupeiformes in bank-solidified sites were significantly higher than in sites with natural shorelines (Fig.4).The top ten most differentially abundant OTUs between bank-solidified and natural shoreline sites were mainlyHypophthalmichthyssp andSinipercasp.
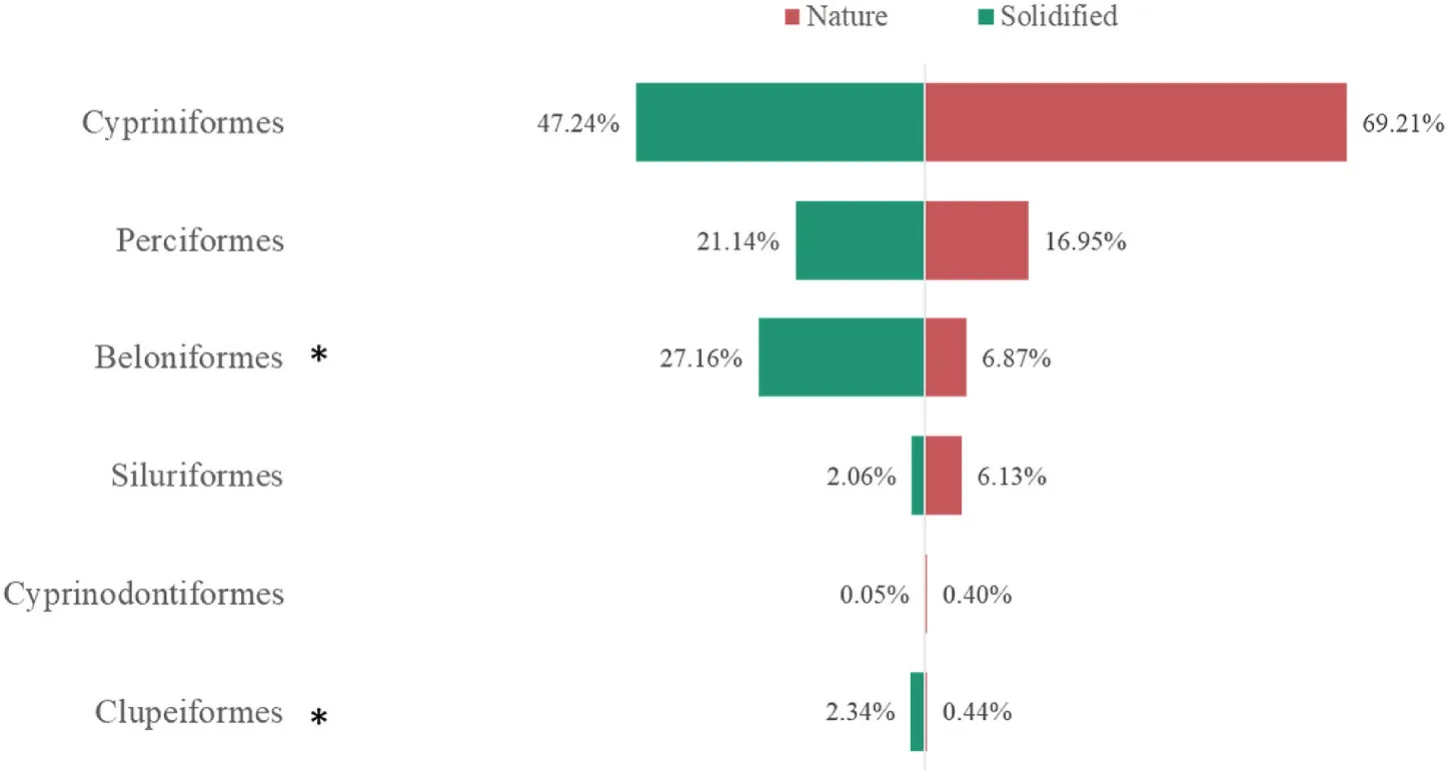
Fig.4.Significant differentially abundant OTUs in the all detected phyla.The phyla label with * indicate a significant difference between bank solidified and nonsolidified sites.

Table 4PERMANOVA on the bank solidified and non-solidified data sets.
Although FCC diversity decreased with the increasing human population, this relationship was not consistent for all teleost.Nonparametric correlation tests demonstrated that most species were not sensitive to changes in the human population.Human populationassociated variables significantly influenced only nine species, and all of them were from the Cyprinidae family (Table 5).
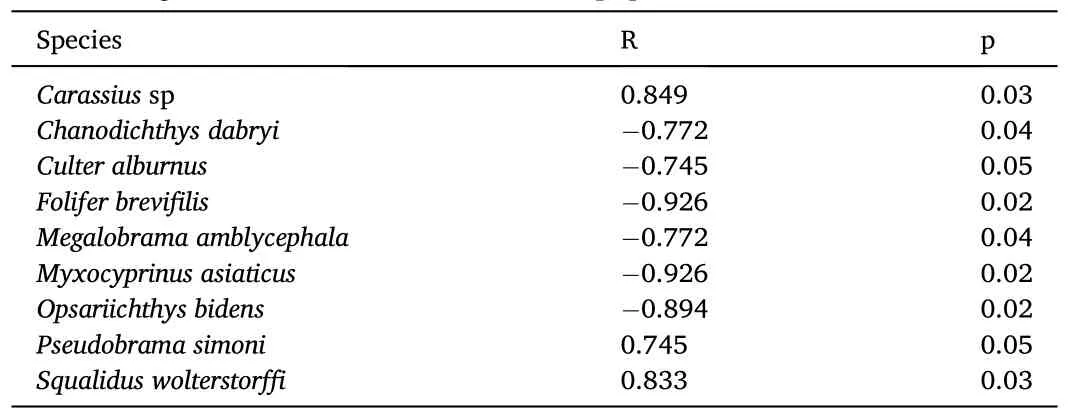
Table 5Fish that significant correlated with human population-associated variables.
3.3.Invasive species and interactions with native species
In total, 102 OTUs (6.14%) were identified from invasive species,and they were from Clariidae, Loricariidae and Poeciliidae families.Among these OTUs, 96 were annotated asClarias gariepinus, a cultured species of African origin.The abundances of OTUs from Loricariidae and Poeciliidae were rare in all sample sites (<1%).Although OTUs annotated asC.gariepinuswere detected at all sites, these OTUs’ frequency was also less than 1% except at the XJHY site (2.55%).Despite the uncommon occurrence ofC.gariepinusat all sites, interaction network analyses showed that this species was highly correlated with 16 native species (i.e., significant correlation coefficients greater than 0.8 or less than -0.8 (P <0.05).The most-influenced native species wasPseudobrama simoni.Among the correlated native species,Sinipercasp was the only carnivorous species related to invasive species, and others were almost omnivorous species (Fig.5).

Fig.5.Native and invasive species interactions network.The circles represent the relative OTUs abundance of each species.
4.Discussion
Human population centers are often concentrated along with freshwater bodies and rivers, and, in particular, the intensity of human impacts on rivers is likely to be particularly severe.The environmental impact of high human density causes adverse effects on the fish composition and community structure (Ghasemi et al., 2015).In the present study, the ichthyofauna in urban sections of Xiang River was compared using eDNA as a case study to investigate the influences of human-induced disturbances on the fish community.
4.1.Assessment of eDNA metabarcoding detection
The region in12sand16sgenes are widely adopted as common DNA barcode regions in research (Liu et al., 2016; Bylemans et al., 2018; Stat et al., 2019).The primers used in this study were the optimal candidate among 14 primer-pairs (Bylemans et al., 2018).They were all successfully amplified in the tested species, indicating that it was promising to detect ichthyofauna in the Xiang River.However, the specificity of the primer was low in this study, with more than half of the amplifications sourced from non-target species.Moreover, because there was no definitive barcoding database, about 22% of sequences could not be annotated (Fig.S1).
The barcoding databases for teleost taxonomic groups were mainly the NCBI database (nr) (https://www.ncbi.nlm.nih.gov/genbank/) and Barcode of Life Database (BOLD) (http://www.boldsystems.org/).Due to the limited length of the barcoding sequences, some OTUs could not be annotated to the species level (eg.Bedotiasp) or were labeled within hybrid species-complexes (e.g.H.nobilis×H.molitrix).Higherspecificity primers and longer reads could be developed for 3rd generation sequencing platforms to alleviate this issue.A local and customized barcoding database based on fish assemblages in the Xiang River would offer a higher resolution for local species identifications.
Although there were limitations on the species identification, the FCC variations were calculated by the frequency of the OTUs, as well as the diversity indexes, and the results could be robust to interpret the community variations without the annotation information.Therefore,FCC variations detected the eDNA metabarcoding in this study supported that the promising prospect of eDNA technique in aquaculture and conservation biological studies.
4.2.Factors that influence fish community biodiversity
Xiang River is about 948 km in length and 94,721 km2coverage and is the chief river of the Dongting Lake drainage system of the middle Yangtze (Li et al., 2018; Zeng et al., 2015).As the largest river in Hunan Province, 60% of the human population lives along the River, and contributes about 3/4 of the provincial GDP (Liu et al., 2010).Anthropogenic disturbances from various industries and increasing urbanization are increasing in intensity in the Xiang River; these adverse impacts were previously implicated in fish community declines (e.g.(Gao et al.,2019; Hu et al., 2020)).Physical surveys conducted between 2008 and 2010 did not detect 24% (36/147) of fish species detected in 1983, and larva abundance has sharply decreased (Ding et al., 2010).In the present study, only 32 species were detected in the middle and lower Xiang River.Even with some unidentified and unannotated sequences, FCC was very different from the previous physical surveys.In Changsha Section, where the largest cities in Hunan province are located, the FCC diversity was in strong decline.
The industrial waste, agricultural nutrients, and domestic sewage from anthropogenic activities can influence the physicochemical parameters of rivers when the human population rapidly increases and this may, in turn, affect the diversity of fauna (Hassan et al., 2014; N’Zi et al.,2015).In the present study, heavy metal ions (mainly from industry) and TN and TP (mostly from agriculture) were not significantly associated with ichthyofauna community structure (Fig.3).This might be related to a high tolerance of heavy metals relative to the concentration of ions observed at study sites.For example, the 96h LC50of Cd on Silver Carp(H.molitrix) was 1.723 mg/L (Pi et al., 2016), whereas the concentration of Cd across the site (0.072-0.297 μg/L) was far less than its LC50(Table S1).
The human population density was negatively associated with fish diversity, but its impact was not caused by variation in sediment and water physiochemical alternations.Riparian habitat degradation due to human environmental modification of fish habitat can be a primary cause of fish diversity declines (Hermoso et al., 2011; Sass et al., 2017;Colin et al., 2018).Riverbanks are an essential component of river habitat, but in increasingly urbanized environments,flooding and erosion can become serious concerns (Coleman et al., 2011).As a result,concrete structures and impervious surfaces have historically been employed to prevent erosion and stabilize riverbanks.However, such channelization can adversely affect biological communities by altering substrate and degrading water quality with possible eutrophication(Hohensinner et al., 2018; Li et al., 2013).Our findings similarly suggest that bank solidification can have an adverse effect on fish diversity, and it was closely linked to the habitat loss at land-water ecotone after channelization.Additionally, the negative relation between biodiversity and the human population revealed that human-derived stresses (e.g.,lights and noises) were possibly expelled the fish as well (Reid et al.,2019), and it could be one of the reasons that low diversity observed in Changsha Section.
4.3.Relationship between alien and native species
The introduction of exotic species to the river ecosystem is a globally common phenomenon, especially in the rivers that flow through cities(Leuven et al., 2009).Collection records from the 1983 survey do not indicate the presence of exotic species, andGambusia affiniswas the only invasive species detected during 2008-2010 surveys (Cao et al., 2012).However, several ornamental fish (Loricariidae and Poeciliidae) were also detected in this study; we speculate that these exotic species originated as escapees from the aquarium industry.Meanwhile, the construction of dams shifted river habitat to a more controlled, stable, and calm hydrological regime, affording a suitable environment for these exotic fish to establish the populations (Li et al., 2013).The management of introduced ornamental fish remains a global challenge (Collins et al.,2013), and the number of these non-native species may increase in the future.
Clarias gariepinus, an invasive species widely distributed throughout the Xiang River, was a popular aquaculture species approximately 30 years ago but has subsequently faded from the markets.Escapees from aquaculture likely contributed to its widespread invasion of the Xiang River.In this study, higher negative correlations were observed between invasive species and native omnivorous species, and it potentially indicated the occurrence of predation or competition between these two kinds of fish for food resources.Also, the eggs of native species may also be preyed upon by some invasive species (e.g.species in Poeciliidae).Competitive interactions between invasive and native fishes could potentially alter trophic exchange and modify food webs in the ecosystem (Carey & Wahl, 2010; Nagelkerke et al., 2018).Native species are also at risk of hybridization with invasive species.Although it was impossible to examine whether the invasive species detected at our sites have established stable populations, the number of sequences recovered from these species indicated that their presence should not be ignored.
4.4.Consequences of hydropower dam
Since the first hydropower station on the Xiang was constructed in1980, eight dams have subsequently been installed on the main stem of the Xiang River in Hunan Province; three of them are located among Hengyang, Changsha and Yueyang.The successive construction of a cascade of hydropower dams has divided and weakened the continuity and integrity of the river ecosystem (Zhong & Power, 1996; Li et al.,2013), with the original diverse habitats replaced by relatively homogenous reservoir habitats.The dams also represent significant structural barriers that have caused habitat fragmentation for fish and other aquatic organisms (Li et al., 2013; Wozney et al., 2011; Wu, 2003).The hydrological characteristics of the Xiang river have been completely modified after the damming, particularly with respect to slowing water velocities and increasing depth (Cao et al., 2012).In present study, low detection ratios ofCtenopharyngodon idellaandMylopharyngodon piceuswere observed in the Changsha section, and it could be related to the habitat shift by the hydro-constructions (Ding et al., 2010; Li et al.,2013; Wu, 2003).However, no significant differences in FCC observed between the sections were divided by the dams.This did not mean that the dams did not cause the isolations of ichthyofauna populations.The homogeneous FCC might be contributed by the artificial propagation and releasing recovery.
4.5.Artificial propagation and releasing recovery
To protect species of conservation concern and recover wild populations, human-mediated propagation of the four major Chinese carps and a protected species Chinese sucker into the Xiang River has been conducted since the beginning of this century.Since 2001, artificial propagation and releasing of grass carp (C.idella), black carp (M.piceus),silver carp (H.molitrix), bighead carp (H.nobilis), crucian carp (Carassius carassius), common carp (Cyprinus carpio), and Chinese sucker (Myxocyprinus asiaticus) (National 2nd Class Protection) has been employed to preserve ichthyofauna biodiversity in the Xiang River.The significant recovery of these carp populations was reported based on their abundances at different sections (Animal Husbandry & Aquaculture Bureau,2015), and these successes resulted in a close FCC in different sections along the River, which could mitigate the disconnections cause by the dams.
The results in the present study showed that the effectiveness of recovery varied substantially between species.Hypophthalmichthyssp(1.12%-65.30%) were detected at the highest abundance (both in reads and OTUs) in this study.We suspect that the main contributor to this high abundance was the artificial releases due to the low genetic diversity observed for this group (Table S3), which was also supported by microsatellite-based parentage assignment (Animal Husbandry &Aquaculture Bureau, 2015).However, the low frequency (<1%) ofCtenopharyngodon IdellaandMylopharyngodon piceusreflected that both species have failed to establish substantial populations in different sections of the Xiang River.These discrepancies could be explained by habitat preferences, becauseH.nobilisandH.molitrixwere benthopelagic while the other two were demersal (www.fishabe.org).Release and propagation strategies should consider the habitat preferences of different species.
4.6.Conservation implications
Examining the community’s structures among three urban sections of the Xiang River illustrates that the ichthyofauna is significantly disturbed by anthropogenic activities.To mitigate the adverse impacts of urbanization, Xiang River’s conservation management should focus on at least three key aspects: river habitat, human disturbances, and fish populations.Constructions in or near the river should be limited to minimize habitat alterations, and an ecological-friendly riverbank should be considered to replace cement constructions.Secondly, the effect of human disturbances on the ecosystem should be minimized,including the perceiving interference (such as, lights and noise) and introduced exotic species.To recover declining populations of key fish species, artificial propagation should be continued with additional considerations, including increasing the number of native species to maintain community composition and prevent alpha biodiversity loss.The increase of omnivorous species could also assist native species due to their negative relationship with invasive species (Gu et al., 2018).Enlarging gene pools are also important to reduce genetic homogeneity,requiring an increase in the number of dams and sires involved in fingerling production.Integrating these three aspects could protect the ichthyofauna from anthropogenic disturbances and reduce biodiversity loss from stresses due to increasing human populations.
Credit authors contribution statement
CZ and DL contributed to research design; BJ, XL and JY provided logistical support for specimen collection; CZ and YW completed all molecular experiments and analysis; all authors contributed to the writing of the manuscript and the interpretation of the data.
Declaration of competing interest
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, the manuscript entitled, “Impact of anthropogenic activities on changes of ichthyofauna in the middle and lower Xiang River”.
Acknowledgment
The authors are particularly grateful to Dr.Matthew C.Yates, Dr.Dylan J.Fraser and Dr.Chanjun Qu for improving the manuscript.The authors also thank Libo Sheng for the sample collections.This study was supported by the Key projects of Hunan Provincial Education Department (No.19A222), Hunan Provincial Natural Science Foundation(2018JJ3236), National Key Research and Development Program of China (No.2019YFD09006030), and National Natural Science Foundation of China (No.31772832 and 31960254).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.aaf.2021.06.007.
 Aquaculture and Fisheries2022年6期
Aquaculture and Fisheries2022年6期
- Aquaculture and Fisheries的其它文章
- Comparison for ecological economic performance of Chinese sea perch(Lateolabrax Maculatus) under different aquaculture systems
- Environmental variables affecting the gillnet catches and condition of Labiobarbus festivus and Osteochilus hasseltii in northern Malaysia
- Genetic diversity and relatedness inferred from microsatellite loci as a tool for broodstock management of fine flounder Paralichthys adspersus
- Retention of fin clips and fin and operculum punch marks in rainbow trout
- Preliminary data of life history traits of Mormyridae (Actinopterygii:Teleostei) in the Upper Sanaga River, Central Region of Cameroon
- Hybrids production as a potential method to control prolific breeding in tilapia and adaptation to aquaculture climate-induced drought