
Marker-Assisted Breeding of Thermo-Sensitive Genic Male Sterile Line 1892S for Disease Resistance and Submergence Tolerance
2021-01-13 07:30:32LuoYanchangMaTingchenJoanneTeoLuoZhixiangLiZefuYangJianboYinZhongchao
Rice Science 2021年1期
Luo Yanchang, Ma Tingchen, Joanne Teo, Luo Zhixiang, Li Zefu, Yang Jianbo, Yin Zhongchao, 3
Research Paper
Marker-Assisted Breeding of Thermo-Sensitive Genic Male Sterile Line 1892S for Disease Resistance and Submergence Tolerance
Luo Yanchang1, 2, Ma Tingchen2, Joanne Teo1, Luo Zhixiang2, Li Zefu2, Yang Jianbo2, Yin Zhongchao1, 3
(Temasek Life Sciences Laboratory, 1 Research Link, National University of Singapore, Singapore 117604, Republic of Singapore; Key Laboratory of Rice Genetics and Breeding, Rice Research Institute, Anhui Academy of Agricultural Sciences, Hefei 230031, China; Department of Biological Sciences, National University of Singapore, 14 Science Drive 4, Singapore 117543, Republic of Singapore)
Rice line 1892S is an elite thermo-sensitive genic male sterile (TGMS) line for two-line hybrid rice production. However, 1892S is susceptible to rice blast, bacterial blight and submergence. Here we reported the introduction of blastresistance ()gene, bacterial blightgeneand submergence tolerance geneinto 1892S genetic background through backcrossing andmarker-assisted selection.The improved TGMS line 31892S and its hybrids conferred disease resistance to rice blast and bacterial blight, and showed submergence tolerance for over 14 d without significant loss of viability. The sterility- fertility conversion of 31892S was similar to that of 1892S. 31892S and its derived hybrid rice had similar agronomic traits and grain quality with 1892S and the control hybrid rice, respectively.The newly developed 31892S provided an improved TGMS line for two-line hybrid rice production with disease resistance to rice blast and bacterial blight, and submergence tolerance with no yield penalty or change in grain quality.
thermo-sensitive genic male sterility;bacterial blight; rice blast; submergence tolerance; disease resistance; marker-assisted selection
Rice (L.) is a major staple food for approximately one-half of the world’s population. However, rice plantation is frequently affected by biotic and abiotic stresses, which cause severe yield loss and have an adverse impact on food security. Rice blast, caused by the fungus, is the most devastating fungal disease in rice plantation in the world. The fatal leaf and panicle blast can even cause yield loss between 10% and 30% with high incidence of infected crops (Sakthivel et al, 2009). To counteractinfection, rice and its wild species have evolved disease resistance () genes. The rice blastgenein line 75-1-127 was introgressed from the wild species(Liu et al, 2002).shows high and broad-spectrum resistance to 43isolates collected from 13 countries (Liu et al,2002),and has been deployed in rice breeding program (Koide et al, 2011; Luo and Yin, 2013; Khanna et al, 2015; Ni et al, 2015; Luo et al, 2016, 2017). Bacterial blight of rice, caused bypv(), is one of the important bacterial diseases that is prevalent throughout the major rice-growing countries, especially in Asia (Gnanamanickam et al, 1999). Plants infected at the maximum tillering stage could have 50% yield reduction (Mew et al, 1993). Rice and its wild relatives have evolved dominant or recessivegenes that confer disease resistance to bacterial blight.is a dominantgene that provides broad-spectrum resistance tostrains (Ikeda et al, 1990), which has been widely deployed in rice breeding for bacterial blight resistance since late 1990s (Huang et al, 1997; Zhang et al, 2006; Perez et al, 2008; Luo et al, 2012, 2014, 2016).
Flood and drought are two major abiotic stresses that adversely affect rice production in Asian countries.Rice variety FR13A was identified to confer submergence tolerance for up to two weeks at the seedling stage (Xu and Mackill, 1996). A major locusfor submergence tolerance in FR13A has been cloned (Xu et al, 2006). The mechanism offor submergencetolerance is to dampen ethylene production and gibberellic acid responsiveness during submergence, so as to economize carbohydrate reserves and to prolong endurance of rice plants (Xu et al, 2006). Interestingly,also enhances the recovery of plants from drought at the vegetative stage through the reduction of leaf water loss, lipid peroxidation and increase in expression of genes associated with acclimation to dehydration (Fukao et al, 2011). Thelocus from FR13A has been introduced into popular rice varieties, including Swarna, KDML105, Mianhui 725, IR64 and 9311 through marker-assisted selection (Neeraja et al, 2007; Septiningsih et al, 2009; Luo and Yin, 2013; Luo et al, 2016, 2017). The improved rice lines with thelocus are tolerant to submergence and maintain high yield, good grain quality and other elite agronomic properties of the recurrent parents.
The development of hybrid rice, known as the second ‘green revolution’ in China, has contributed significantly to rice production and food security in China for over 40 years (Yuan, 2014). So far, two systems on hybrid rice seed production have been developed and successfully commercialized. The three-line hybrid rice system includes a cytoplasmic male sterile (CMS) line, a maintainer line and a restorer line for a commercial production of rice hybrids. The CMS line is used as a maternal line to cross with the restorer line as a pollen donor for hybrid seed production. The maintainer line maintains the sterility of the CMS line and is used to produce the seeds of the CMS line. The two-line hybrid rice system has only two parental lines, a photoperiod-sensitive genic male sterile (PGMS) line or a thermo-sensitive genic male sterile (TGMS) line as the maternal line and the pollen donor as the paternal line. Unlike a CMS line in three-line hybrid rice system that requires a maintainer line for its propagation, a PGMS or TGMSline can be propagated by self-pollination under permissive conditions of photo and/or temperature combinations. In addition, as the male sterile trait of a PGMS or TGMS line is controlled by a single recessive nuclear gene, any conventional rice varieties can be used as pollen donors to produce F1hybrids. Therefore, the two-line hybrid rice technology simplifies the system for hybrid rice seed production, increases the probability of identifying heterotic hybrid combination, especially in yield and grain quality, and saves the cost on hybrid seed production. Annong S-1 is anTGMS rice line that carries male sterile gene(Deng et al, 1999). Thegene was cloned in 2014 (Zhou et al, 2014). The elite TGMS line1892Salso carries thegene (Zhou et al, 2014). 1892S has been used as the maternal line for more than 25 two-line hybrid rice varieties released in China, which had total plantation area of more than 1.696 million hectares during 2006–2014 (Yang et al, 2016). However, 1892S is susceptible to most of the isolates or strains of rice blast or bacterial blight pathogens (Song et al, 2016). Our unpublished preliminary study also indicated that, like most of rice varieties, 1892S is susceptible to submergence.Here we reported the development of an improved TGMS line by pyramiding,andinto 1892S genetic background for disease resistance to rice blast and bacterial blight and submergence tolerance through backcrossing breeding and marker- assisted selection.
Results
Marker-assisted breeding of 31892S
To introduce and pyramid the,andgenes in 1892S genetic background, 1892S was used as the female to cross with donor line T4105, carrying,andgenes in KDML105 genetic background, followed by backcrossing using 1892S as the recurrent female (Fig. 1). Marker-assisted selection was conducted to select positive F1and backcross plants at each generation (Fig. 1 and Table S1). In summary, 24 F1, 3 BC1F1, 4 BC2F1, 11 BC3F1, 2 BC4F1, 6 BC5F1and 5 BC6F1plants were identified to carry heterozygous alleles at the,andloci (Fig. 1). The 5 BC6F1plants were self-pollinated and produced a BC6F2population with 1 550 individuals. Genotyping analysis demonstrated that one BC6F2plant (BC6F2-945) carried homozygous alleles at the,,andloci (Fig. 1). Subsequently, the BC6F2-945 plant was designated as 31892S. The patterns of molecular markers for the,andloci in 3198S were similar to those in T4105 and the corresponding original donor lines, 75-1-127, IR64- Sub1A and IRBB21 (Fig. 2-A to -C). 31892S also carried homozygousgenes from 1892S (Fig. 2-D) and showed sterile phenotype at restrictive conditions. 31892S and 1892S had similar morphological phenotype and growth duration. 31892S was allowed to set seeds at permissive conditionsin the autumn season in Hefei, Anhui Province or in the winter season in Lingshui Hainan Province, China for further study.

Fig. 1. Marker-assisted breeding of 31892S.
The donor line T4105 is a BC4F2plant containing homozygous,andgenes in KDML105 genetic background. The number of total plants screened and the number of positive plants identified are indicated in each generation. Only one plant, BC6F2-945, was identified from 1 550 BC6F2individuals to contain homozygous alleles at the,,andloci. BC6F2-945 was designated as 31892S.
Disease resistance of 31892S and its F1 hybrids against rice blast and bacterial blight
To evaluate 31892S and its F1hybrids for rice blast resistance, the seedlings of rice lines 75-1-127, 1892S, 31892S, RH003, and F1hybrids 1892S/RH003 and 31892S/RH003 were challenged with two fieldisolates, 11-17-1-2-2 and 11-3-1-1-2. 75-1-127 and 31892S were resistant to the two isolates, whereas 1892S was susceptible to bothisolates (Fig. 3-A and -B). As for the F1hybrids, the control 1892S/ RH003 was susceptible to 11-17-1-2-2, whereas 31892S/ RH003 was resistant to theisolate (Fig. 3-A). As RH003, the paternal line of the F1hybrids, was resistant to 11-3-1-1-2, and both 1892S/RH003 and 31892S/RH003 F1hybrids conferred disease resistance to theisolate (Fig. 3-B). 31892S also showed disease resistance to rice blast at the maturity stage in field, while neck blast infection was observed in 1892S plants (Fig. 3-C and -D). The results collectively demonstrated that thegene in 31892S and its F1hybrid 31892S/RH003 conferred disease resistance to rice blast.
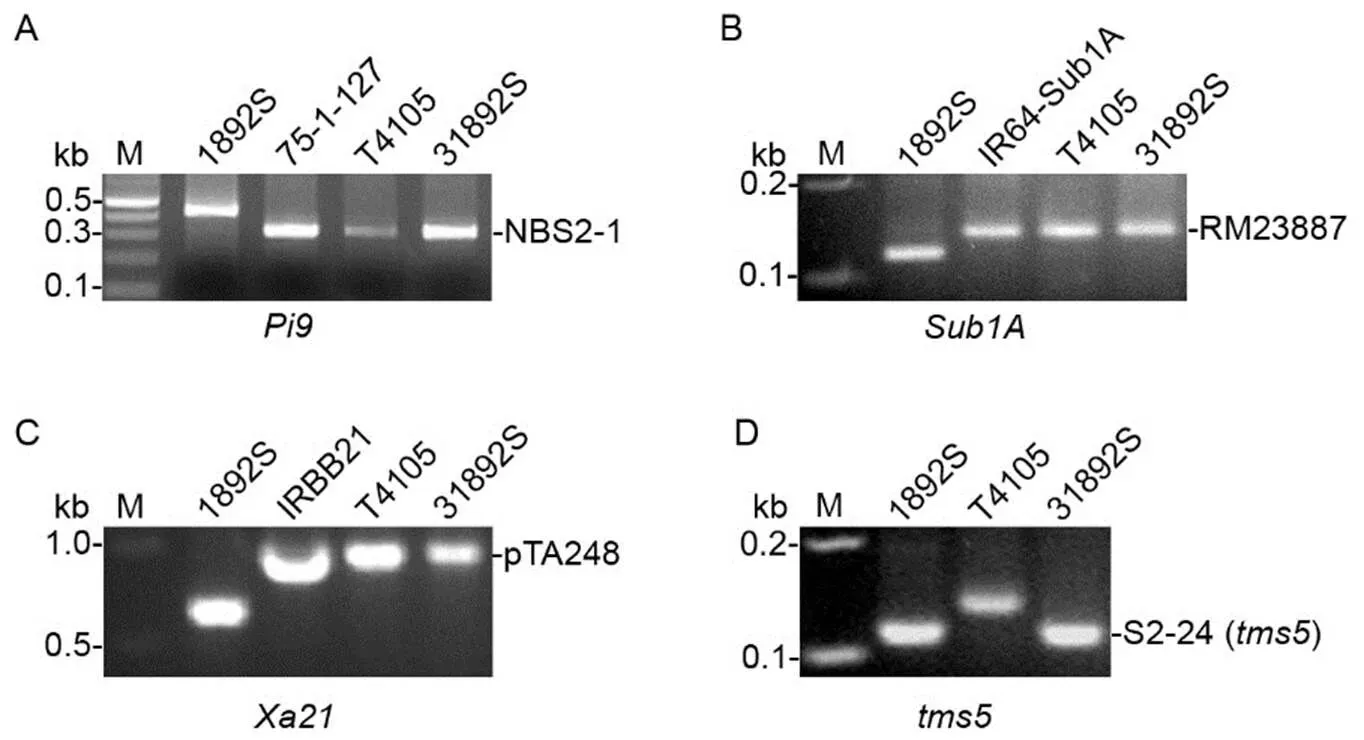
Fig. 2. Genotyping of molecular markers at the selected loci in 31892S.
A, Genotyping of sequence-tagged site (STS) marker NBS2-1 at thegene. 75-1-127 is the original donor line of thegene.B, Genotyping of simple sequence repeat (SSR) marker RM23887 closely linked to thegene. IR64-Sub1A is the original donor line of thegene.C, Genotyping of STS marker pTA248 co-segregated with thegene. IRBB21 is the original donor line of thegene.D, Genotyping of SSR marker S2-24 closely linked to thegene in 1892S.M, Marker.
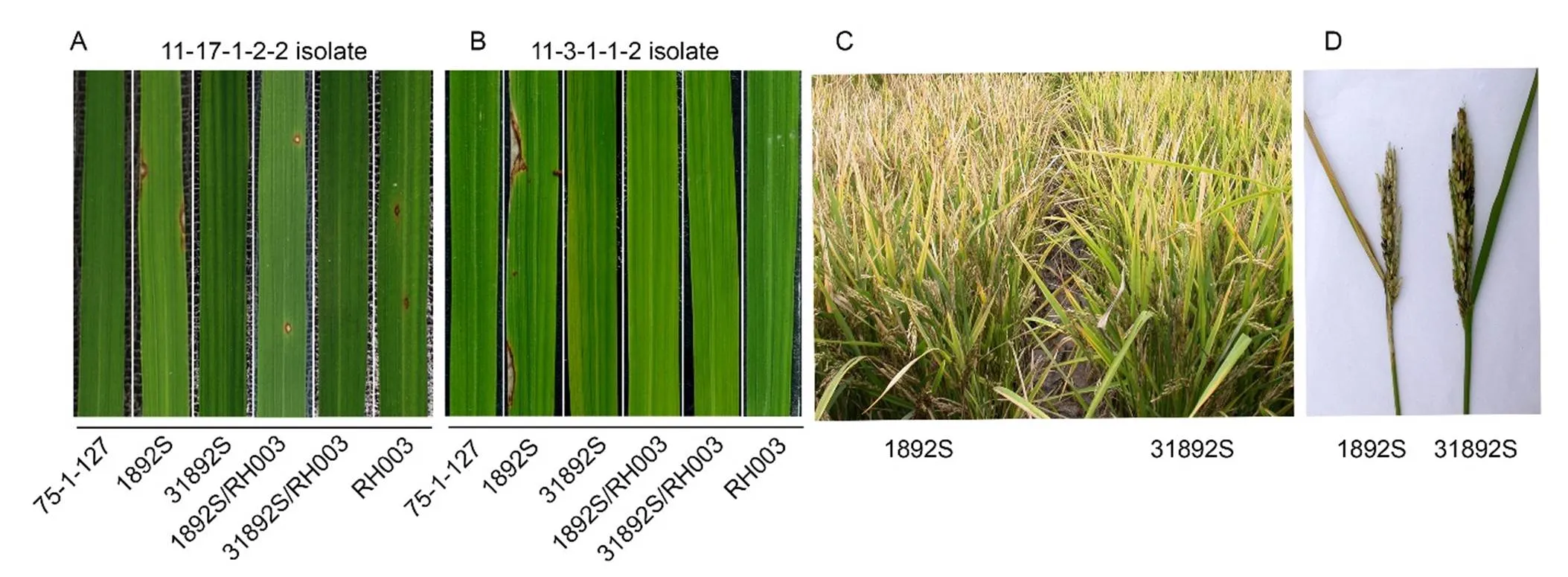
Fig. 3. Disease evaluation of rice lines for resistance to rice blast.
A and B, Disease evaluation of rice lines to rice blast. Two-week-old rice seedlings were inoculated withisolates 11-17-1-2-2 (A) and 11-3-1-1-2 (B). The images were taken at 7 d after blast inoculation.C, Phenotypes of 1892S and 31892S in field. Image was taken at 30 d after heading in the field trial in Hefei, Anhui Province, China, in the summer season of 2017.D, A typical flag leaf and panicle of 1892S with neck blast infection and a healthy flag leaf and panicle of 31892S with no blast infection.75-1-127,introgressive line; 1892S, Thermo-sensitive genic male sterile (TGMS) line; 31892S, Improved TGMS line in 1892S genetic background; RH003, Paternal line used for hybrid F1production; 1892SS/RH003, F1hybrid derived from the cross between 1892SS and RH003; 31892S/RH003, F1hybrid derived from the cross between 31892S and RH003.
31892S and its F1hybrids were tested for bacterial blight resistance with 24strains collected from 9 countries. The originaldonor line IRBB21 conferred resistance or moderate resistance to 21 strains. 1892S was susceptible or moderately susceptible to 18 strains (Table 1). RH003 was susceptible or moderately susceptible to 20 strains. The control F1hybrid 1892S/RH003 was susceptible or moderately susceptible to 20 strains. Collectively, among the 24strains tested, 18 strains could cause susceptible or moderately susceptible phenotypes on 1892S, RH003 and 1892S/RH003. The improved line 31892S was resistant or moderately resistant to 17 of the 18 1892S-compatiblestrains, and the improved F1hybrid 31892S/RH003 conferred resistance or moderate resistance to 23 of the 24strains tested, including the 18 compatiblestrains on 1892S, RH003 and 1892S/RH003 (Table 1). The results demonstrated that thegene in 31892S and its F1hybrid 31892S/RH003 provided broad spectrum resistance tostrains.
Submergence tolerance of 31892S and its F1 hybrids
Fourteen-day-old seedlings of IR64-Sub1A, 1892S, 31892S, RH003, 1892S/RH003 and 31892S/RH003 were tested for submergence tolerance (Fig. 4-A). After 14 d of complete submergence, 1892S, RH003 and 1892S/RH003 plants turned yellow, while IR64- Sub1A, 31892S and 31892S/RH003 plants maintained green or light green (Fig. 4-B). Plant viability was evaluated based on the emergence and growth of new leaves over 7 d of recovery from the 14 d of complete submergence. Most of the IR64-Sub1A (85.6% ± 5.1%), 31892S (85.6% ± 13.5%) and 31892S/RH003 (78.9% ± 12.6%) plants survived from the 14 d of submergence, while most of the 1892S (4.4% ± 1.9%), RH003 (8.9% ± 1.9%) and 1892S/RH003 (6.7% ± 0.0%) plants died (Fig. 4-C and -D). The results indicated that thegene in 31892S and its F1hybrids conferred tolerance to submergence.
Sterility-fertility alteration of 31892S
Sterility-fertility alteration of spikelets on panicles of 31892S and 1892S plants was examined with bagged panicles during the summer season of 2017 in Hefei, China. The spikelets on panicles of 31892S were completely sterile when the panicles were bagged on 27 July 2017 to 7 September 2017 (Fig. 5-A). Spikelet fertility was partially resumed when the panicles were bagged on 10–19 September 2017 (Fig. 5-A). Therefore, the stable period of complete male sterility of 31892S was at least 43 d, which was similar to the bagged panicles of 1892S (Fig. 5-B). After sterility-fertility alteration, the spikelet fertility of the bagged panicles of 31892S ranged from 11.1% ± 7.2% to 24.7% ± 11.1%, which was similar to that of 1892S (from 11.2% ± 6.5% to 21.8% ± 7.1%) (Fig. 5-A and -B). No pollen grain was observed in the spikelets of 31892S or 1892S before sterility-fertility alteration, and no filled seed was set on the panicles when they were grown under restrictive conditions (Fig. 5-C to -E). Fertile pollen grains were observed in the panicles of 31892S and 1892S after sterility-fertility alteration as the temperature dropped to permissive levels (Fig. 5-F and -G). Both 31892S and 1892S had normal spikelet fertility and similar seed-setting rate when grown in the winter season in Lingshui, China (Fig. 5-H). In summary, the fertility-sterility alteration of 31892S was similar to that of 1892S.

Fig. 4. Evaluation of rice lines for submergence tolerance.
A,Phenotypes of plants at 14 d after germination and before submergence treatment. B, Phenotypes of plants after 14 d of submergence treatment. C, Phenotypes of plants after 7 d of recovery from submergence treatment. D, Viability of plants after 7 d of recovery from submergence treatment. The data represent Mean ± SD (= 3, about 30 plants for each line in each treatment). The different capital letters indicate significant difference among rice lines at the 0.01 level according to the Duncan’s multiple range tests. 1,donor line IR64-Sub1A (genotype:); 2, 1892S; 3, 31892S; 4, RH003; 5, 1892S/RH003; 6, 31892S/RH003.
Agronomic traits and grain quality of 31892S and its F1 hybrids
Three field trials were conducted for 1892S and 31892S with one in the summer season of 2017 in Hefei, China,and two in the winter seasons of 2015/2016 and 2016/ 2017 in Lingshui, China. Meanwhile, two field trials were conducted for the F1hybrids, 1892S/RH003 and 31892S/RH003, in the summer seasons of 2016 and 2017 in Hefei, China. Important agronomic traits were collected from the plants in the field trials, and grain quality was measured. 31892S and 1892S had similar growth duration, plant height, productive panicle number per plant, panicle length andtotal number of spikelets per panicle in all field trials under either restrictive or permissive conditions (Table 2). There was no significant difference in spikelet fertility, 1000-grain weight and yield when they were grown under permissive conditions(Table 2). The results indicated that 31892S and 31892S/RH003 had similar important agronomic traits to 1892S and 1892S/RH003, respectively (Table 2).
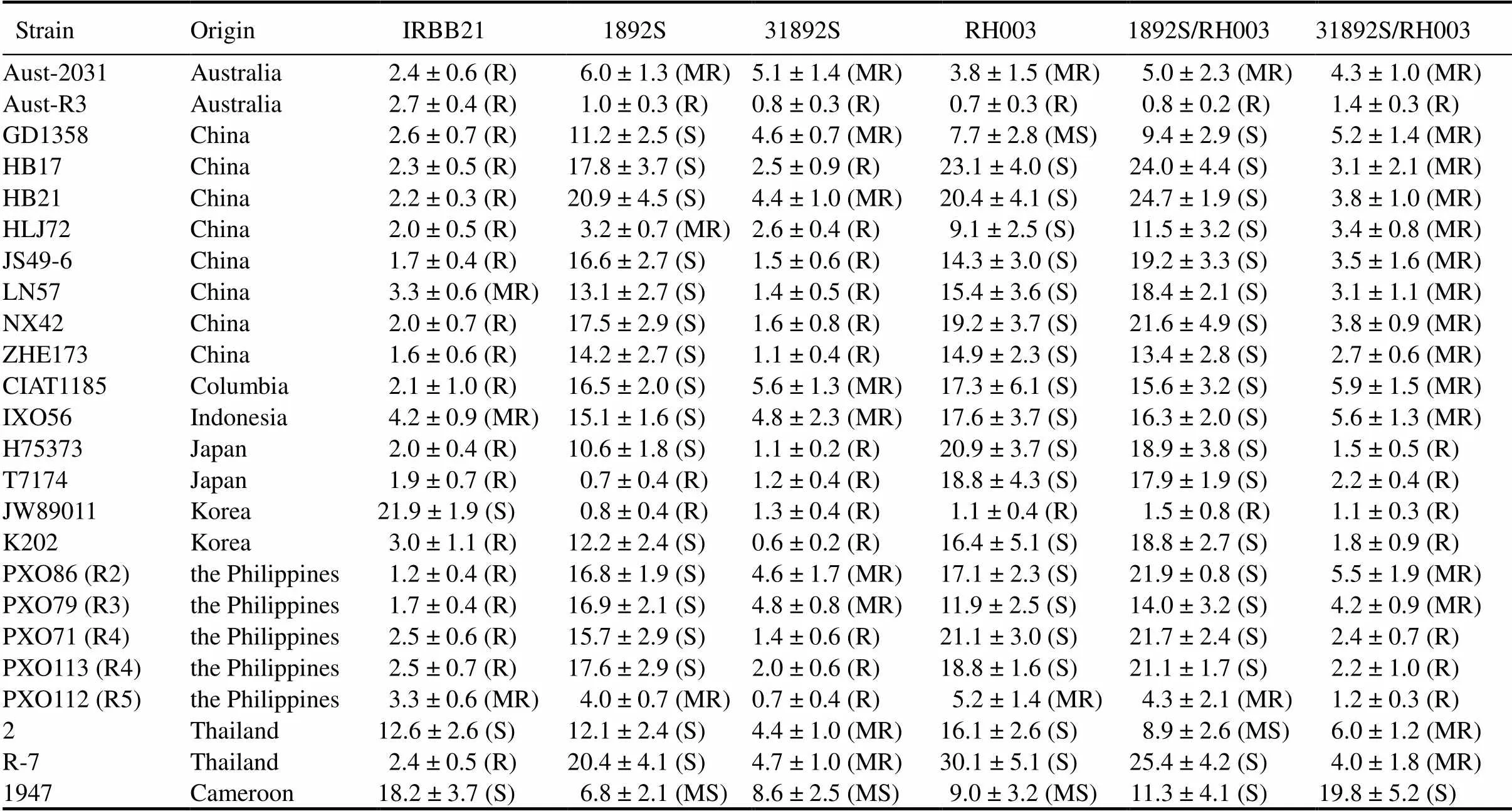
Table 1. Disease evaluation of IRBB21, 1892S, 31892S and F1 hybrids to rice bacterial blight.
R, Resistance (lesion length ≤ 3.0 cm); MR, Moderate resistance (3.0 cm < lesion length ≤ 6.0 cm); MS, Moderately susceptible (6.0 cm < lesion length ≤ 9.0 cm); S, Susceptible (lesion length > 9.0 cm). The lesion length of bacterial blight is shown in Mean ± SD (= 15).
The grain quality of TGMS lines and hybrid rice (F2grains) were measured. Both 31892S and 1892Swere characterized as medium grain rice based on ratio of length to width (Table 3). They had very low degree of chalkiness (DC) and low amylose content (AC) (Table 3). They were also classified as soft rice with lowgel consistency (GC) and intermediate alkali spreading value (ASV) (Table 3). The results indicated that 31892S retains similar grain quality to 1892S. In fact, the quality of the F2grains harvested from the F1hybrids is more important than that of the parental lines as they are consumed by people rather than being used as the seeds for reproduction. F2grains of both 31892S/RH003 and 1892S/RH003were characterized as medium grain rice (Table 3). They had similar DC and intermediate AC, and were also classified as soft rice with similar GC and ASV (Table 3). The results demonstrated that both 31982S and its derived hybrid rice had similar grain quality to 1892S and the control hybrid rice, respectively.
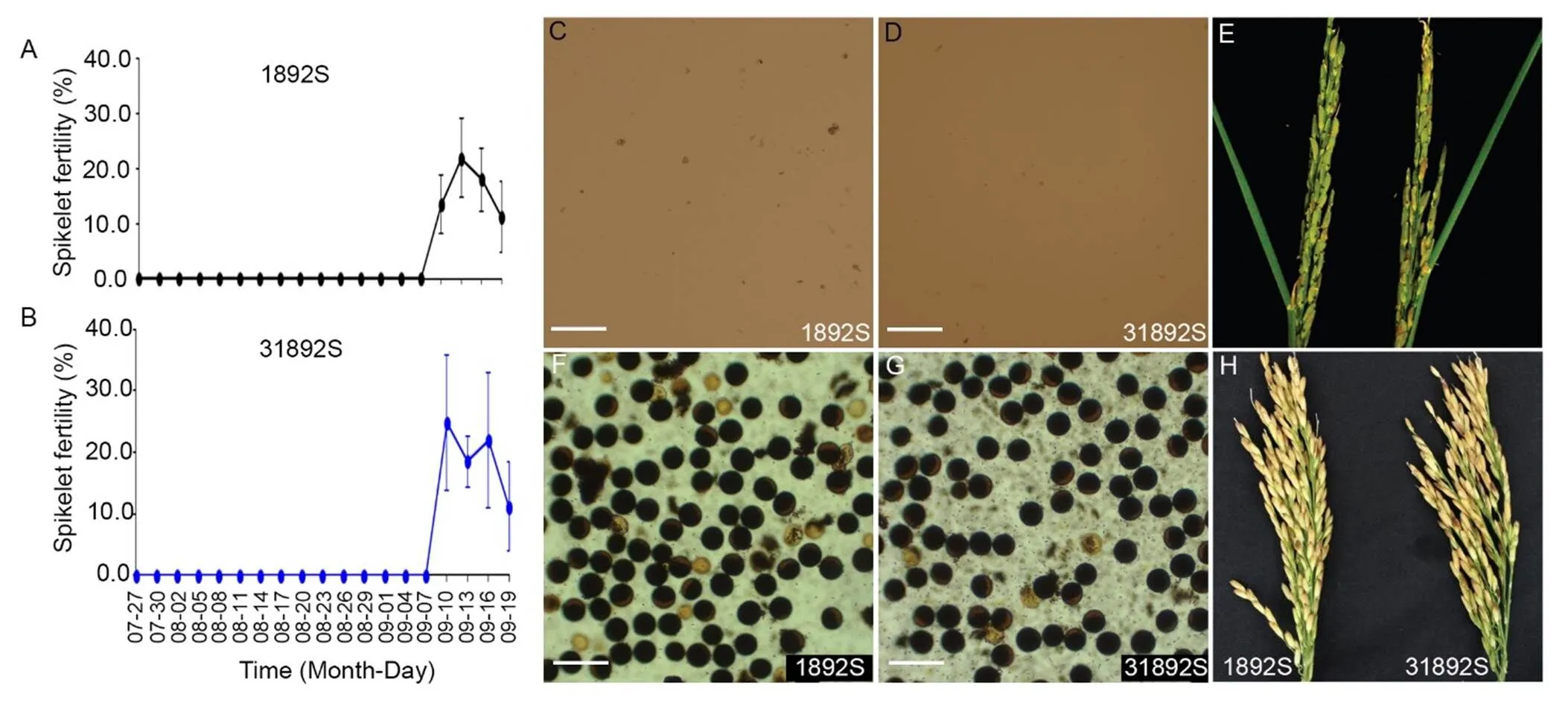
Fig. 5. Sterility-fertility conversion of 1892S and 31892S.
A and B, Spikelet fertility of bagged panicles on 1892S (A) and 31892S (B) plants at different bagged time in 2017.C and D, Pollen grains of 1892S (C) and 31892S (D) plants grown under restrictive conditions with panicles flowering on 14 August, 2017. E, Panicles of 1892S and 31892S plants grown under restrictive conditions in Hefei, China with panicles flowering on 14 August 2017. F and G, Pollen grains of 1892S (C) and 31892S (D) after sterility-fertility alteration with panicles flowering on 18 September 2017 in Hefei, China. H, Panicles of 1892S and 31892S plants grown under permissive conditions in the winter season of 2016/2017 in Lingshui, China. In C, D, F and G, Pollen grains were stained with 1% I2-KI solution. Scale Bars, 100 μm.
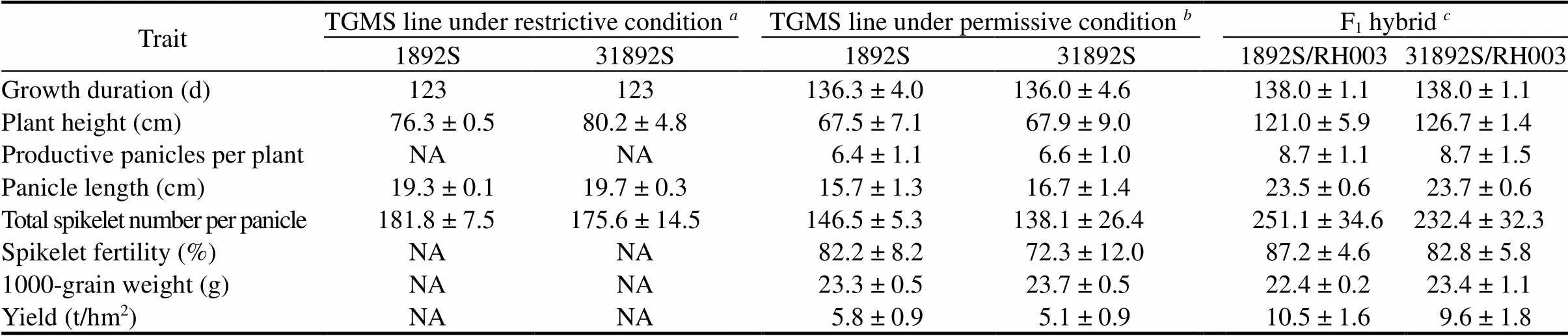
Table 2. Agronomic traits of 1892S, 31892S and their F1 hybrids.
Field trial for thermo-sensitive genic male sterile (TGMS) lines was conducted in Hefei, China, in the summer season of 2017 (= 3).Two field trials for TGMS lines were conducted in Lingshui, China, in the winter seasons of 2015/2016 and 2016/2017 (= 6).Two field trials for hybrid rice plantation were conducted in Hefei, China, in the summer seasons of 2016 and 2017 (= 6).Data represent Mean ± SD. Statistical analysis for-value was performed in Microsoft Office Excel 2007 using a two-tailedtest for paired samples, and there were no significant difference in all.NA, Not available.
Discussion
Hybrid rice has been successfully used for commercial rice production in China for over 40 years. However, the hybrid F1production and plantation are still affected by biotic and abiotic stresses. The utilization of natural resistance in parental lines is still an efficient, economical and sustainable way to combat against the biotic and abiotic stresses. Previously, we have introduced and pyramided the,andgenes in WH725 and 9311 genetic backgrounds and developed two elite paternal lines for two- and/or three-line hybrid rice production (Luo et al, 2016, 2017).In this study, we successfully introduced and pyramided the,andgenes in 1892S genetic background and developed an improved TGMS line 31892S for two-line hybrid rice. 31892S and its F1hybrids conferred disease resistance to rice blast and bacterial blight and provided submergence tolerance. 1892S, an elite TGMS line, has been used as female to produce more than 25 commercial F1hybrids, which have been widely grown in midstream and downstream Yangze Valley (Yang et al, 2016). The development of 31892S will provide enhanced disease resistance to rice blast and bacterial blight and submergence tolerance in two-line hybrid rice production.
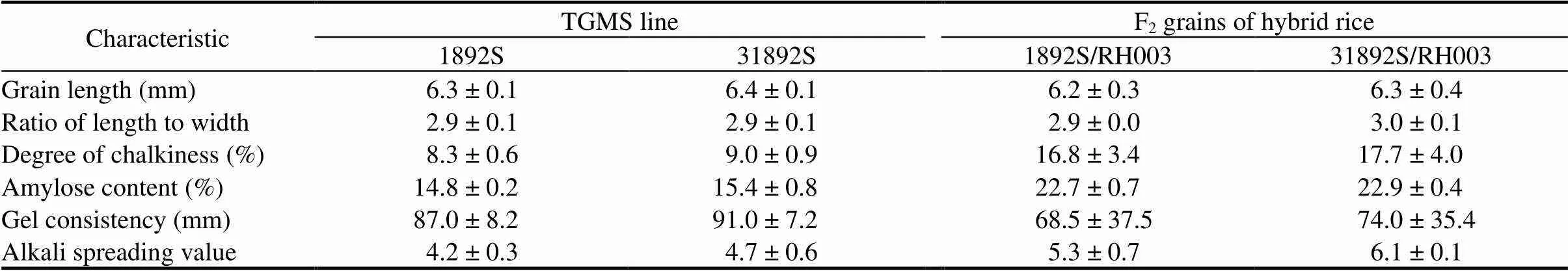
Table 3. Grain quality of 1892S, 31892S and F2 grains of hybrid rice.
The evaluation of grain rice quality of 1892S and 31892S was done with rice seeds from the field trials in the winter seasons of 2015/2016 and 2016/2017 in Lingshui, China. The evaluation of grain quality of hybrid rice was done with rice seeds collected from the field trials in the summer seasons of 2016 and 2017 in Hefei, China. TGMS, Thermo-sensitive genic male sterile.
Backcrossing is a traditional but still efficient way for the introgression of gene(s) from a donor line into a recurrent female line. However, if several genes from different donor lines need to be introduced and pyramided into a single recurrent female line, co-current backcrossing has to be conducted in each generation before gene pyramiding at BC4F1or higher backcrossinggenerations (Luo and Yin, 2013; Luo et al, 2016, 2017).Marker-assisted selection can provide precise selectionwithout doing phenotypic selection. However, co-current backcrossing and subsequent gene pyramiding are still labour-intensive and time-consuming. In this study, T4105, which carries the,andgenes in KDML105 genetic background, was selected as the single donor line for the three genes. Marker-assisted selection of the,andloci was conducted in a single medium-size backcrossing population at each generation from BC1F1to BC6F1. A large population was only generated at the BC6F2generation in order to screen for individual plants that contained homozygous allele at the,andloci. The breeding strategy saved at least one generation of time at the gene pyramiding stage. Most importantly, 31892S and its F1hybrids showed resistance to rice blast and bacterial blight and provided submergence tolerance.
Undesirable genes, linked with a selected gene or locus, which is also known as linkage drag (Brinkman and Frey, 1977), is always a concern in marker- assisted breeding. Backcrossing and marker-assisted selection can reduce the size of introgression window more rapidly and more efficiently. The originaldonor line 75-1-127 is a BC3offspring, derived from the wild cross betweencv. IR31917-45-3-2 (2= 24, AA genome) andAcc. 101141 (2=48, BBCC genome), followed by backcrossing using the cultivar as the recurrent female (Amante-Bordeos et al, 1992). IRBB21 is a near-isogenic line of thegene in IR24 genetic background, which was generated through intensive backcrossing after thelocus was introgressed from the African wild rice(2= 24, AA genome) to cultivated rice (Khush et al, 1990). Thelocus was originally identified in the Indian landrace FR13A (Mackill et al, 1993). IR64-Sub1A is a BC3F2line of thegene in IR64 genetic background (Septiningsih et al, 2009). T4105, the common donor line for the,andgenes, is a BC4F2plant with the three genes pyramided in KDML105 genetic background(Luo and Yin, 2013). Taken together, for all the three genes in this study, at least 15 generations of crossing and/or backcrossing have been conducted withvarieties as the female or recurrent female after they were introgressed from the wild species or the native landrace. 31892S and its F1hybrid 31892S/RH003 had similar agronomic traits and grain quality to 1892S and the control F1hybrid 1892S/RH003, respectively. No pleiotropic phenotype, yield penalty or change in grain quality was observed. The results indicated that linkage drag did not occur during the introducing and pyramiding of the,andgenes in 1892S genetic background. Similar results were also observed in our previous work on introducing and pyramiding the three genes in WH725 and 9311 genetic backgrounds (Luo et al, 2016, 2017). It is estimated that 31892S may harbour narrow introgression windows at the,andloci derived from,and FR13A, respectively.
Methods
Rice materials
1892S is an elite TGMS line carryinggene. T4105 was a sister line of T5105 in KDML105 genetic background with thegene from 75-1-127, thegene from IR64-Sub1A and thegene originally from IRBB21 (Luo and Yin, 2013). RH003 is a paternal line for F1hybrid production.
PCR-based molecular markers
The molecular markers were NBS2-1 for thegene (Luo et al, 2016), RM23887 for thelocus (Neeraja et al, 2007), S2-24 for thelocus (Xu et al, 2011; Zhou et al, 2014) and pTA248 for thelocus (Chunwongse et al, 1993). The nature of the molecular markers and their DNA primers are summarized in Table S1. The molecular markers were amplified by PCR on a BIO-RAD T100 programmable thermal cycler (Life Science Research, USA). The PCR reaction mixture of 20 μL consisted of < 200 ng of rice genomic DNA, 0.15 mmol/L of each primer, 10 μL of 2× PCR GoTaq? Green Master Mix (Promega, USA). Template DNA was denatured at 94 oC for 3 min, followed by 35 cycles of PCR amplification with the following parameters: 30 s of denaturation at 94 oC; 30 s of primer annealing at 55 oC for markers pTA248, RM23887 and S2-24, and 60 oC for marker NBS2-1; 30 s of primer extension for RM23887, S2-24 and NBS2-1, and 90 s of primer extension for pTA248 at 72 oC. Finally, the reaction mixture was maintained at 72 oC for 5 min before completion. The amplified products were electrophoretically resolved on 1.5% agarose gel for pTA248 and NBS2-1, and 3.0% agarose gel for RM23887 and S2-24 in 1× Tris-acetate-EDTA buffer.
M. oryzae isolates and blast inoculation
Fieldisolates 11-17-1-2-2 and 11-3-1-1-2 were collected from rice fields in Anhui Province, China. The isolates were grown on prune agar medium with 40 mL prune juice, 5 g/L lactose, 5 g/L sucrose, 1 g/L yeast extract, 20 g/L agar and pH 6.5 at 28 oC in darkness for 7 d and at 26 oC under light for 4 d. For blast inoculation, two-week-old rice plants were inoculated with a spore suspension at the density of 1 × 105spores/mL. The inoculated plants were placed in darkness in a dew chamber for 24 h at 25 oC and 90% of humidity. The inoculated plants were then transferred to a growth chamber at 25 oC–28 oC and grown under a 12 h/12 h (light/dark) photoperiod and 90% humidity. The disease reaction was scored at 7 d after inoculation.
Xoo strains and bacterial blight inoculation
Twenty-fourstrains collected from nine countries were used for bacterial blight inoculation.strains were grown on peptone-sucrose agar (PSA) medium (10 g/L peptone, 10 g/L sucrose, 1 g/L glutamic acid, 16 g/L agar and pH 7.0) at 28 oC for 2 d. The bacterial cells were collected and suspended in sterile water at an optical density of 0.5 (600). Fully expanded leaves of 6-week-old rice plants were inoculated withstrains by the leaf clipping method (Kauffman et al, 1973). The lesion length was measured at 14 d after inoculation.
Evaluation of rice for submergence tolerance
Tests of rice plants for submergence tolerance were conducted in an open water tank and repeated for three times. In each experiment, 20 plants were tested for each rice line. Fourteen- day-old plants grown in pot soil were completely submerged in water. The submergence-treated plants were allowed to recover for 7 d before scored for viability. The plant survival was indicated by having minimum growth of a new green leaf (Luo et al, 2016). The statistical analysis was carried out by the method of new multiple range test (Duncan, 1955).
Dynamic observation of fertility expression in the field
Tests on fertility-sterility alteration of TGMS lines were conducted in 2017 in Hefei, China. One hundred seeds for each TGMS line were sown at 10-day intervals from 21 April to 11 July 2017. Thirty uniform and healthy rice seedlings at the five-leaf stage were transplanted in fields at a raw spacing of 20.0 cm × 20.0 cm. Ten panicles from each TGMS line were bagged for self-pollination at 3-day interval during the heading period from 27 July to 19 September 2017. Spikelet fertility of the bagged panicles was scored at 25 d after heading. Pollen grains from top five florets of primary panicles were stained with 1% I2-KI solution and examined under a ZEISS Axioplan Upright microscope.
Design of field trials and collection of important agronomic traits
Field trials for the TGMS lines were conducted in Lingshui, China, in the winter seasons of 2015/2016 and 2016/2017 and in Hefei, China in the summer season of 2017. Field trials for the F1hybrids were conducted in Hefei, China, in the summer seasons of 2016 and 2017. For each rice line, three randomly selected plots were planted with each at 12 m2. Major agronomic traits were collected as described previously (Luo et al, 2016).-value of statistical analysis for each trait was calculated according to a two-tailed-test for paired samples.
Evaluation of grain quality
The characteristics of rice grain quality, including grain length, ratio of length/width, degree of chalkiness, amylose content, gel consistency and alkali spreading value (ASV), were measured according to the method described previously (Cruz and Khush, 2000).
Acknowledgements
The authors thank Zhou Youzhi for his help in field trials, Bai Yisong for TGMS line 1892S and Zhang Aifang forisolates. This research is supported by Temasek Foundation, Temasek Life Sciences Laboratory Innovation Fund and the National Research Foundation, Prime Minister’s Office, Singapore, on the research programme of the Disruptive and Sustainable Technology for Agricultural Precision.
SUPPLEMENTAL DATA
The following material is available in the online version of this article at http://www.sciencedirect.com/science/ journal/16726308; http://www.ricescience.org.
Table S1. Molecular markers for genes used in this study.
Amante-Bordeos A, Sitch L A, Nelson R, Dalmacio R D, Oliva N P, Aswidinnoor H, Leung H. 1992. Transfer of bacterial blight and blast resistance from the tetraploid wild riceto cultivated rice,., 84: 345–354.
Brinkman M A, Frey K J. 1977. Yield-component analysis of oat isolines that produce different grain yields., 17(1): 165–168.
Chunwongse J, Martin G B, Tanksley S D. 1993. Pre-germination genotypic screening using PCR amplification of half-seeds., 86(6): 694–698.
Cruz N D, Khush G S. 2000. Rice grain quality evaluation procedures.: Singh R K, Singh U S, Khush G S. Aromatic Rices. New Delhi, India: Oxford and IBH Publishing Co. Pvt. Ltd: 24–36.
Deng H F, Shu F B, Yuan D Y. 1999. An overview of research and utilization of Annong S-1., 14(3): 1–3. (in Chinese)
Duncan D B. 1955. Multiple range and multipletests., 11: 1–41.
Fukao T, Yeung E, Bailey-Serres J. 2011. The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice., 23(1): 412–427.
Gnanamanickam S S, Priyadarisini V B, Narayanan N N, Vasudevan P, Kavitha S. 1999. An overview of bacterial blight disease of rice and strategies for its management., 77(11): 1435–1444.
Huang N, Angeles E R, Domingo J, Magpantay G, Singh S, Zhang G, Kumaravadivel N, Bennett J, Khush G S. 1997. Pyramiding of bacterial blight resistance genes in rice: Marker-assisted selection using RFLP and PCR., 95(3): 313–320.
Ikeda R, Khush G S, Tabien R E. 1990. A new resistance gene to bacterial blight derived from., 40(S1): 280–281.
Kauffman H E, Reddy A P K, Hsieh S P Y, Merca S D. 1973. An improved technique for evaluating resistance of rice varieties to., 57(6): 537–541.
Khanna A, Sharma V, Ellur R K, Shikari A B, Gopala Krishnan S, Singh U D, Prakash G, Sharma T R, Rathour R, Variar M, Prashanthi S K, Nagarajan M, Vinod K K, Bhowmick P K, Singh N K, Prabhu K V, Singh B D, Singh A K. 2015. Development and evaluation of near-isogenic lines for major blast resistance gene(s) in Basmati rice., 128(7): 1243–1259.
Khush K S, Esperanza B, Ogawa T. 1990. A new gene for resistance to bacterial blight from.., 7: 121–122.
Koide Y, Ebron L A, Kato H, Tsunematsu H, Telebanco-Yanoria M J, Kobayashi N, Yokoo M, Maruyama S, Imbe T, Fukuta Y. 2011. A set of near-isogenic lines for blast resistance genes with an-type rainfed lowland elite rice (L) genetic background., 123(1): 19–27.
Liu G, Lu G, Zeng L, Wang G L. 2002. Two broad-spectrum blast resistance genes,(t) and(t), are physically linked on rice chromosome 6., 267(4): 472–480.
Luo Y C, Ma T C, Zhang A F, Ong K H, Li Z F, Yang J B, Yin Z C. 2016. Marker-assisted breeding of the rice restorer line Wanhui 6725 for disease resistance, submergence tolerance and aromatic fragrance., 9(1): 66.
Luo Y C, Ma T C, Zhang A F, Ong K H, Luo Z X, Li Z F, Yang J B, Yin Z C. 2017. Marker-assisted breeding of Chinese elite rice cultivar 9311 for disease resistance to rice blast and bacterial blight and tolerance to submergence., 37(8): 106.
Luo Y C, Sangha J S, Wang S H, Li Z F, Yang J B, Yin Z C. 2012. Marker-assisted breeding of,andin the restorer lines of hybrid rice for broad-spectrum and enhanced disease resistance to bacterial blight., 30: 1601–1610.
Luo Y C, Yin Z C. 2013. Marker-assisted breeding of Thai fragrance rice for semi-dwarf phenotype, submergence tolerance and disease resistance to rice blast and bacterial blight., 32: 709–721.
Luo Y C, Zakaria S, Basyah B, Ma T C, Li Z F, Yang J B, Yin Z C. 2014. Marker-assisted breeding of Indonesia local rice variety Siputeh for semi-dwarf phonetype, good grain quality and disease resistance to bacterial blight., 7(1): 33.
Mackill D J, Amante M M, Vergara B S, Sarkarung S. 1993. Improved semidwarf rice lines with tolerance to submergence of seedlings., 33(4): 749–753.
Mew T W, Alvarez A M, Leach J E, Swings J. 1993. Focus on bacterial blight of rice., 77(1): 5–12.
Neeraja C N, Maghirang-Rodriguez R, Pamplona A, Heuer S, Collard B C Y, Septiningsih E M, Vergara G, Sanchez D, Xu K, Ismail A M, Mackill D J. 2007. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars., 115(6): 767–776.
Ni D H, Song F S, Ni J L, Zhang A F, Wang C L, Zhao K J, Yang Y C, Wei P C, Yang J B, Li L. 2015. Marker-assisted selection of two-line hybrid rice for disease resistance to rice blast and bacterial blight., 184: 1–8.
Perez L M, Redona E D, Mendioro M S, Cruz C M V, Leung H. 2008. Introgression of,andfor resistance to bacterial blight in thermosensitive genetic male sterile rice (L.) for the development of two-line hybrids., 164(3): 627–636.
Sakthivel K, Sundaram R M, Rani N S, Balachandran S M, Neeraja C N. 2009. Genetic and molecular basis of fragrance in rice., 27(4): 468–473.
Septiningsih E M, Pamplona A M, Sanchez D L, Neeraja C N, Vergara G V, Heuer S, Ismail A M, Mackill D J. 2009. Development of submergence-tolerant rice cultivars: thelocus and beyond., 103: 151–160.
Song F S, Ni J L, Zhang A F, Wang C L, Zhao K J, Li L, Ni D H, Yang J B. 2016. Molecular improvement of male sterile line 1892S for resistance to rice blast and bacterial blight., 14(6): 1507–1515. (in Chinese with English abstract)
Xu J J, Wang B H, Wu Y H, Du P N, Wang J, Wang M, Yi C D, Gu M H, Liang G H. 2011. Fine mapping and candidate gene analysis of, the photoperiod-thermo-sensitive genic male sterile gene in rice (L.)., 122(2): 365–372.
Xu K N, Mackill D J. 1996. A major locus for submergence tolerance mapped on rice chromosome 9., 2(3): 219–224.
Xu K N, Xu X, Fukao T, Canlas P, Maghirang-Rodriguez R, Heuer S, Ismail A M, Bailey-Serres J, Ronald P C, Mackill D J. 2006.is an ethylene-response-factor-like gene that confers submergence tolerance to rice., 442: 705–708.
Yang L S, Bai Y S, Qian Z. 2016. Promotion and application of male sterile line 1892S breeding series resistant combination., 44(8): 63–66. (in Chinese with English abstract)
Yuan L P. 2014. Development of hybrid rice to ensure food security., 21(1): 1–2.
Zhang J, Li X, Jiang G, Xu Y, He Y. 2006. Pyramiding ofandfor the improvement of disease resistance to bacterial blight in hybrid rice., 125(6): 600–605.
Zhou H, Zhou M, Yang Y Z, Li J, Zhu L Y, Jiang D G, Dong J F, Liu Q J, Gu L F, Zhou L Y, Feng M J, Qin P, Hu X C, Song C L, Shi J F, Song X W, Ni E D, Wu X J, Deng Q Y, Liu Z L, Chen M S, Liu Y G, Cao X F, Zhuang C X. 2014. RNase ZS1processesL40mRNAs and controls thermosensitive genic male sterility in rice., 5: 4884.
Copyright ? 2021, China National Rice Research Institute. Hosting by Elsevier B V
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/)
Peer review under responsibility of China National Rice Research Institute
http://dx.doi.org/10.1016/j.rsci.2020.11.010
29 January 2020;
9 May 2020
Yin Zhongchao(yinzc@tll.org.sg)
(Managing Editor: Wang Caihong)

- Rice Science的其它文章
- Different Hypotheses for Resistance Loss of Rice Varieties to Magnaporthe oryzae
- Development of Heat Tolerant Two-Line Hybrid Rice Restorer Line Carrying Dominant Locus of OsHTAS
- Identification of QTLs for Cadmium Tolerance During Seedling Stage and Validation of qCDSL1 in Rice
- Genetic and Geographic Patterns of Duplicate DPL Genes Causing Genetic Incompatibility Within Rice: Implications for Multiple Domestication Events in Rice
- Physiochemical Properties of Resistant Starch and Its Enhancement Approaches in Rice
- Anticancer Activities of Plant Secondary Metabolites: Rice Callus Suspension Culture as aNew Paradigm