
Comparative Skin Histology of Frogs Reveals High-elevation Adaptation of the Tibetan Nanorana parkeri
2019-06-26 03:03:30ChunhuaYANGTingtingFUXinqiangLANYunZHANGLotannaMicahNNEJIRobertMURPHYYanboSUNJingCHE
Asian Herpetological Research 2019年2期
Chunhua YANG, Tingting FU, Xinqiang LAN, Yun ZHANG, Lotanna Micah NNEJI, Robert W. MURPHY,5, Yanbo SUN*, Jing CHE,6*
1 State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, Chinese Academy of Sciences, 650223 Kunming, China
2 Institute of Physical Science and Information Technology, Anhui University, 230601 Hefei, China
3 Kunming College of Life Science, University of Chinese Academy of Sciences, Kunming 650204, China
4 Key Laboratory of Animal Models and Human Disease Mechanisms, The Chinese Academy of Sciences and Yunnan Province, Kunming Institute of Zoology, Chinese Academy of Sciences, 650223 Kunming, China
5 Centre for Biodiversity and Conservation Biology, Royal Ontario Museum, 100 Queen's Park, Toronto, Ont., M5S 2C6,Canada
6 Center for Excellence in Animal Evolution and Genetics, Chinese Academy of Sciences, 650223 Kunming, China
Abstract Adaptations to extreme environmental conditions are intriguing. Animal skin, which directly interacts with external environment, plays diverse and important roles in adaptive evolution. The thin and bare skin of amphibians is sensitive to external environmental conditions and, thus, it facilitates investigations into adaptations for living in extreme environments. Herein, we compare the structures of skin in four anuran species living at elevations ranging from 100 m to 4500 m to assess phenotypic innovations in the skin of Nanorana parkeri, which lives at extremely high elevations. Analyses reveal similar basic skin structures, but N. parkeri differs from the other species by having more epidermal capillaries and granular glands, which correlate highly with responses to hypoxia and/or ultraviolet (UV)radiation. Further intraspecific comparisons from frogs taken at ~4500 m and ~2900 m reveal that all of the changes are fixed. Changes occurring only in the higher elevation population, such as possessing more skin pigments, may represent local adaptations to coldness and/or UV radiation. These results provide a morphological basis for understanding further the molecular adaptations of these frogs.
Keywords Nanorana parkeri, skin structure, phenotypic innovation, high-elevation adaptation
1. Introduction
Environmental adaptation is an intriguing topic in evolutionary biology (Cheviron and Brumfield, 2009;Cheviron et al., 2014; Storz et al., 2010). This includes topics such as environment-genotype interactions and the role(s) natural selection plays in driving phenotypic changes (Denver, 2009; Fusco and Minelli,2010). The skin of animals forms the first protective barrier to external stimuli as it interacts directly with the surrounding environment (Huang et al., 2016).Accordingly, skin always experiences strong selection pressures at physiological and/or morphological levels when an organism faces a new or changed environment.Skin innovations can benefit survival greatly. For example, variations in human skin color are adaptive in response to ultraviolet (UV) radiation (Jablonski and Chaplin, 2010). Thus, investigations into basic phenotypes of skin in varying environments, such as high versus low elevations, may provide insights into how organisms adapt to their environment.
Unlike most other vertebrates whose skin is covered by either scales or hair, the thin and bare skin of amphibians plays an important role in respiration (Tattersall, 2007).This condition causes amphibians to be more sensitive to their external environment than most other vertebrates.Some species, like Nanorana parkeri, an endemic frog of the Qinghai-Tibet Plateau (QTP), can live up to 5000 m a.s.l. (Che and Wang, 2016) where challenges such as hypoxia (Hassanin et al., 2009), strong UV radiation (Norsang et al., 2011) and cold temperatures(Ma and Lu, 2010) occur. Thus, this frog is an excellent model for investigating adaptations of skin to extreme environmental conditions.
Previous studies have examined some morphological and physiological differences of skin among frogs living at different elevations. For instance, the skin of Rana kukunoris, a species living at ~3867 m, was shown to have more pigment-bearing chromatophores and cutaneous glands than R. chaochiaoensis from 1935 m and R. omeimontis from 308 m) (Mi and Liao, 2016).Because chromatophores may serve to absorb heat and synthesize melanin (Del Bino and Bernerd, 2013; Liang and Wang, 2004), the increase in density is advantageous for coping with both coldness and UV radiation. Further,cutaneous glands secrete several antimicrobial peptides that resist pathogens and UV radiation (Brizzi et al.,2002; Delfino et al., 2015; Melzer et al., 2013; Rota et al., 2017). Their proliferation likely constitutes another adaptation to living at high elevations. Similarly, middlehigh elevation odoriferous frogs like Odorrana andersonii(~2000 m) can secrete more skin antioxidant peptides than lower elevation O. wuchuanensis (~700 m) (Yang et al., 2016). Further, high-elevation Scutiger have abundant capillaries, which potentially promote gaseous exchange under hypoxia (Cao et al., 2011). These studies suggest that increases in epidermal capillaries, chromatophores and cutaneous glands in the stratum spongiosum might constitute adaptations of these frogs to high elevations.
Phenotypic changes occur in different groups of frogs,yet no study explores these potential adaptations within one high-elevation species and few analyses exist on adaptations of a species versus local adaptations. Further,even fewer studies compare morphological phenotypes among different high-elevation frogs (e.g., N. parkeri versus R. kukunoris), which to some extent limits our understanding of skin innovations of frogs. Within a species, like N. parkeri whose distribution spans from~2900 m to ~5000 m (Che and Wang, 2016), variation may exist as a function of elevation.
To address these issues, we analyze four closely related anuran species, that occur on a range of elevations(Che and Wang, 2016; Che et al., 2010), and investigate phenotypic changes along their phylogeny. Analyses include N. parkeri (“High” group collected at ~4500 m;“Low” collected at ~2900 m), N. medogensis (~2800 m),N. phrynoides (~1800 m), and Quasipaa spinosa (~170 m). Key to the analysis, N. parkeri is one of known highest elevation anurans and yet it occupies a wide span of elevations (Che and Wang, 2016) and High emerged most recently (Wang et al., 2018). Finally, potential molecular adaptations acquired by N. parkeri have been investigated on a genome-wide scale (Sun et al., 2018;Sun et al., 2015; Wang et al., 2018), and this provides an opportunity to evaluate the genotype-phenotype association in skin evolution.
2. Materials and Methods
2.1. SamplingApproval of the project (BBCJ-2014-001)was obtained from Kunming Institute of Zoology (KIZ)and sampling followed the animal use protocols approved by the Animal Care and Ethics Committee of KIZ. All species (Table 1) were collected during their reproductive periods. Three adult males per species/population were sampled, and their snout-vent length (SVL) were measured using a digital Vernier caliper to ensure they were of similar age. The mean SVL and standard deviation for all species were then calculated (Table 1).
2.2. Skin histologyTo examine the basic skin structures,we used the Hematoxylin-eosin (HE) staining procedures(Yang et al., 2016). Dorsal skin was excised near the foramen magnum of euthanized individuals. These tissues were fixed in 10% buffered formalin for about 24 hours and subsequently preserved in 70% ethanol under ambient conditions. Tissues were then embedded in paraffin.Histological sections with 6 μm thickness were cut on positively charged slides, which were then stained with the HE solution.
2.3. Image digitization and statistical analysisImages of each histological section were captured using a Leica DM4000B microscope (Leica, Heidelberg, Germany)at ×100 or ×200 magnification. Skin pigment content(mean integrated optical density, MIOD; Greenberg et al.,1995), skin thickness (in millimeters) and the number of epidermal capillary vessels and glands were calculated with Image Pro-Plus? (IPP) version 6.0 (http://www.mediacy.com/imageproplus). Statistically significant differences in skin structures were evaluated using t-tests (two-tailed) at the P < 0.05 level of significance as implemented in Origin 2017 (https://www.originlab.com/).
3. Results
3.1. Similar basic skin structuresThe frogs had similar basic skin structures composed of epidermis and dermis.Generally, the epidermis was a pluristratified epithelium,primarily composed of keratinocytes. The dermis was divided into two parts, the stratum spongiosum and stratum compactum, which were mainly comprised of loosely and densely compacted cells, respectively (Figure 1). Pigment-bearing chromatophores and capillaries occurred in the stratum spongiosum. Collagen fibers in the stratum compactum occurred in bundles that ran parallel to the body surface. Typical of amphibians, the frogs had mucous and granular glands in the stratum spongiosum(Figure 1). Notwithstanding, some differences existed among the samples. For example, N. parkeri lacked a calcified layer at the junction between the stratum spongiosum and stratum compactum (Figure 1).
3.2. Epidermis layer and epidermal capillary vesselsWe compared all frogs with respect to the thickness of the epidermis layer and the number of epidermal capillaries,because these factors potentially affect the respiratory function of the skin (Brizzi et al., 2002; Tattersall, 2007).For example, a thinner epidermis and more epidermal capillaries were thought to enhance skin respiration(Liang and Wang, 2004; Tattersall, 2007). These attributes varied in our frogs with elevation, particularly at High.Individuals at High had a thinner epidermis layer (P< 0.001, Figure 2A) than other populations. However,individuals at Low and low-elevation species had comparable thicknesses (Figure 2A). The significant difference between High and Low populations constituted a localized change that occurred after the divergence of High from Low.
With respect to the number of epidermal capillaries,both High and Low populations had significantly more epidermal capillary vessels than the other species (P <0.001, Figure 2B), which seemed to be an innovation in N. parkeri. Notwithstanding, High also had significantly more capillaries than Low, and this likely represented another localized adaptation.
3.3. Pigment-bearing chromatophoresPigmentbearing chromatophores were imbedded in the stratum spongiosum (Figure 1) and fewer skin pigments occurred in the epidermis and around granular glands of lower elevation species. In comparison, High had significantly higher pigment content (MIOD) in the both dermal and epidermal layers than did the other three species (P <0.001; Figure 2C). Even when compared with Low, High had remarkably higher pigment content. However, Low and the other three species did not exhibit prominent differences.
3.4. Granular and mucous glandsMucous and granular glands were the most common forms of glands; granular glands were shown to secret diverse peptides important for defense (Toledo and Jared, 1995). Our results showed that High and Low have significantly more granular glands but less mucous glands than the other studied species (P < 0.001, Figure 2D and E).
4. Discussion
4.1. Potential mechanisms for the loss of calcified layer in N. parkeriThe calcified layer is thought to have originated from residues of ancestral dermal bones, and it is comprised of calcium phosphate, proteoglycan and mucopolysaccharide. Its functions may include regulation of calcium level and assisting in substance exchange(Toledo and Jared, 1993).
A comparison of the loss of the calcified layer in N.parkeri to other high-elevation frogs serves to investigate loss as a potential adaptation. Scutiger boulengeri, S.glandulatus and S. muliensis, which live at about 3000 m have a calcified layer, but it consists of discontinuous slender fragments (Cao et al., 2011). The calcified layer exists in lower elevation R. chaochiaoensis (~1900 m)and R. omeimontis (~300 m) (Mi and Liao, 2016). Thus,the loss of the calcified layer in N. parkeri may be an adaptation and the species of Scutiger may be eliminating the layer. Alternatively, the loss may be a random event,or, potentially adaptation to other environmental factors.Future functional and developmental studies can shed light on these possibilities.
4.2. Fixed adaptation of epidermal capillary vessels and cutaneous glandsFrogs from High have more epidermal capillaries than other species, even more than Low (Figure 2B). The inferred population history of N. parkeri (Wang et al., 2018), and its phylogenetic relationships (Che and Wang, 2016; Che et al., 2010)suggest that the number of epidermal capillaries is an important target of natural selection because the species experienced at least two increases in population size during its evolution: after speciation; and following the split of High and Low. The continuum suggests that angiogenesis is important for coping with the highelevation environments. In support of this, angiogenesisrelated genes were shown to have experienced rapid evolution after speciation (Sun et al., 2018). Hence, a thinner epidermis and an increase in capillaries appear to be adaptations to living at high-elevations.
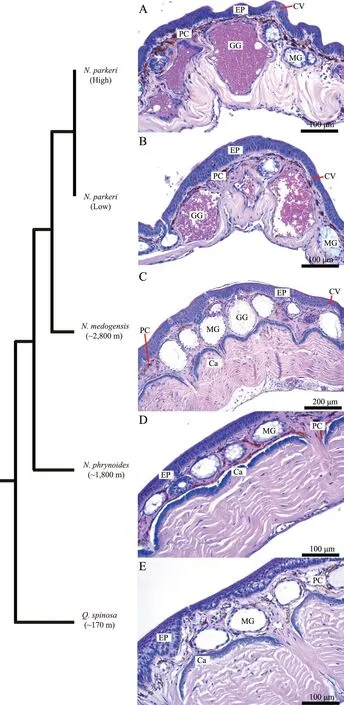
Figure 1 Basic structures of anuran skin. Phylogenetic relationships of selected species (branch lengths are not patristic distances)modified from Che et al. (2010). Cross-sections of the dorsal skin of (A) Nanorana parkeri (High) (×200), (B) N. parkeri (Low) (×200),(C) N. medogensis (×100), (D) N. phrynoides (×200) and (E) Quasipaa spinosa (×200). Note: Ca, calcified layer; CV, capillary vessel; EP,epidermis; GG, granular gland; MG, mucous gland; PC, pigment cell.
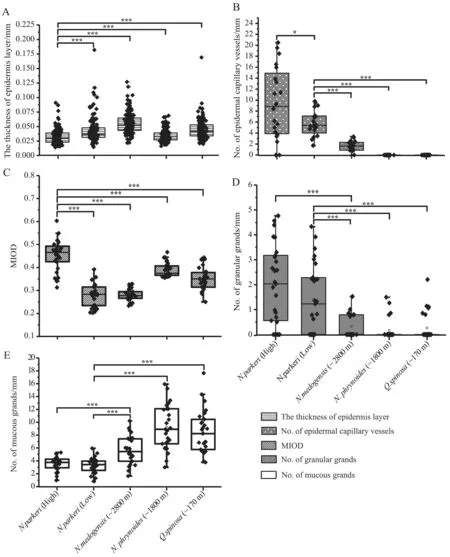
Figure 2 Morphological changes in the dorsal skin of Nanorana parkeri, N. medogensis, N. phrynoides and Quasipaa spinosa. Bar plots show (A) thickness of epidermis layer of the anuran species; (B) total number of epidermal capillary vessels; (C) mean integrated optical density (MIOD = IOD/skin pigment total area of HE staining), which is a measure of total content of the skin pigment; (D) total number of granular glands; and (E) total number of mucous glands. (Two-tailed t-tests: *** P < 0.001, * 0.01< P < 0.05).
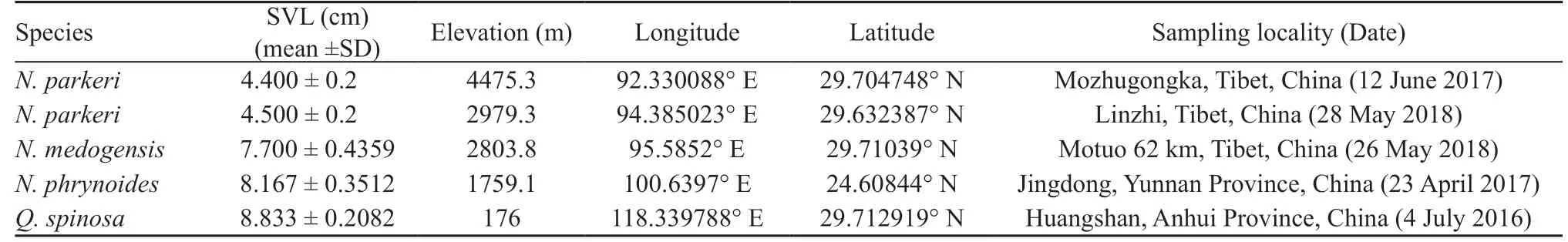
Table 1 Species information including date and locality of sampling, and snout-vent length (SVL).
Mucous and granular glands, the most common forms of glands in amphibian skin, serve important defense functions. Nanorana parkeri has significantly more granular glands than the other species, and this modification may be a defense adaptation. Because most previous studies on cutaneous glands did not compare species at different elevations (Cao et al., 2011; Jia L.L. et al., 2013; Liang and Wang, 2004), comparative analyses can only use R. kukunoris (~3800 m), R.chaochiaoensis (~1900 m) and R. omeimontis (~300 m).Rana kukunoris and R. chaochiaoensis also have more granular glands than do the other species of Rana (Mi and Liao, 2016). Both High and Low have significantly more granular glands than other studied species (Figure 2D).Thus, we hypothesize that this increase of granular glands is an adaptation to its living at high elevations. Given the lack of data on mucous glands of other anurans, further study is necessary to test this hypothesis.
4.3. Local adaptations to high elevations enhanced by pigment-bearing chromatophoresNanorana parkeri has significantly more chromatophores in the both dermal and epidermal layers than do the other studied frogs. This phenotype was observed in R. kukunoris (Mi and Liao,2016). Because cellular pigments are advantageous for absorbing heat and synthesizing melanin (Kim et al.,2005; Pan et al., 2012), greater pigment content may be a local adaptation acquired independently by high elevation species, which experience both coldness and stronger UV radiation.
Species inhabiting different elevations differ in their skin structures. The Tibetan frog appears to have evolved numerous phenotypic changes in skin structure.These potential adaptations may include having a thinner epidermis layer, more epidermal capillaries and more pigment-bearing chromatophores. Other highelevation frogs appear to have acquired these changes independently (Cao et al., 2011; Liang and Wang, 2004;Mi and Liao, 2016) and future genome-level study can determine if this occurred via convergent or parallel evolution. Notwithstanding, N. phrynoides (1800 m)has a thinner epidermis and higher pigment content than some frogs living at higher elevations (N. medogensis and Low). High-elevation adaptation starts at about 1800 m(Sun et al., 2018); abundant phenotypic and genotypic modifications occur in frogs at this elevation. Thus, both the thickness of the epidermis and pigment content closely associate with living at high elevations. Several studies have focused on potential molecular adaptations of the Tibetan frogs (Sun et al., 2018; Yang et al., 2012) and our study provides an important resource for associating the genotype and phenotype in a frog, as well as enhancing an understanding of evolutionary strategies. Functional laboratory experiments of the effects of UV radiation,hypoxia and cold temperature on the structure of frogs'skin remain necessary to complete painting of the picture on anuran adaptations to extreme environments.
AcknowledgmentsWe thank Yi ZHANG, Baolin ZHANG, Xue MI, and Song HUANG for their assistance during the sample collection and laboratory procedures.This study was supported by the National Natural Science Foundation of China Grant (31671326 and 31871275).Jing CHE and Yanbo SUN are supported by the Youth Innovation Promotion Association, Chinese Academy of Science, China.
 Asian Herpetological Research2019年2期
Asian Herpetological Research2019年2期
- Asian Herpetological Research的其它文章
- Evaluating the Importance of Environmental Variables on Spatial Distribution of Caspian cobra Naja oxiana (Eichwald,1831) in Iran
- On the Generic Taxonomy of Opisthotropis balteata (Cope, 1895)(Squamata: Colubridae: Natricinae): Taxonomic Revision of Two Natricine Genera
- Molecular Phylogeny and Evolution of Two Rhacophorus Species Endemic to Mainland Japan
- Genetic Structure of the Red-spotted Tokay Gecko, Gekko gecko(Linnaeus, 1758) (Squamata: Gekkonidae) from Mainland Southeast Asia
- Appendix 1 Specimens examined.