
Seaweed nutraceuticals and their therapeutic role in disease prevention
2019-05-26 01:54:26AirmiGnesnUmTiwriGurvRjuri
食品科學(xué)與人類健康(英文) 2019年3期
Airmi R. Gnesn, Um Tiwri, Gurv Rjuri
a School of Applied Sciences, College of Engineering, Science and Technology, Fiji National University, Fiji
b School of Science, Institute of Technology Sligo, Ballinode, Co. Sligo, Ireland
c School of Agriculture and Food Science, University College Dublin, Lyons Research Farm, Celbridge, Co. Kildare, Ireland
Keywords:
A B S T R A C T
1. Introduction
The usage of seaweeds as traditional food and complementary medicine was recorded in early archaeological data of ten thousand years back [1]. They are traditionally consumed in many Asian countries like China, Indonesia, Philippines, South Korea,North Korea, Japan and Malaysia for centuries, however, the culinary use of seaweed started in Japan and China. Recently, they have attained more reputation in western countries and now are widely utilized as food in the USA, South American and European countries due to their functional properties and the introduction of Asian cuisine [1]. Nowadays, the popularity of seaweed has become a more versatile form of a food ingredient that can be incorporated directly or indirectly in the preparation of foods and beverages. Due to its functionality seaweed and its products are of particular importance in the food industry, as components in fertilizers, animal feed supplements and additives for functional foods [2]. They are low caloric food but rich in vitamins, minerals and essential trace elements, polyunsaturated fatty acids, bioactive metabolites, proteins, polysaccharides and dietary fibres. Apart from regular consumption, many studies advocated the health benefits of seaweed supplementation alongside a regular diet. Regular consumption of seaweed effectively reduced the depressive symptoms among pregnant Japanese women as well decreased the risk of suicide in adults [3,4]. Frequent consumption of dietary seaweeds decreased the risk of diabetes mellitus in the Korean population [5].They are also utilised as hydrocolloids, as emulsifier and as gelling agents in various food product preparations [3].
In addition to benefits of regular consumption of seaweed in diet, the medicinal properties of seaweed bioactive have been historically recognized. For example, seaweeds are used for treatment and or for prevention of goitre, which is caused by the lack of iodine in the diet [6]. Several studies have shown various remedial effects of algal species against non-communicable diseases such as inflammation, obesity, diabetes, hypertension and viral infections [7]. A clinical study indicated that regular consumption of Undaria seaweed can effectively minimise the risk of breast cancer in women [8], while an oral administration of seaweed extracts (Fucus vesiculosus, Macrocystis pyrifera and Laminaria japonica) with zinc, manganese and vitamin B6,potentially decreased osteoarthritis symptoms in a mixed population [9]. Besides its extensive medicinal properties, seaweeds are recognized for its antioxidant capacities and bioactive polyphenolic compounds [10]. Studies have also demonstrated potential roles in HIV protection, primarily linked with compounds present in algae such as phlorotannins, sulfated polysaccharides, certain diterpenes and lectins [11]. Additionally, cancer prevention and metabolic syndrome (METS) associated with obesity, cardiovascular diseases, diabetes, and chronic inflammation are key attributes of algae in relation to human health and wellness [12,14].Furthermore, both fermentable and insoluble dietary fibres are fundamental components found in macroalgae helps to ameliorate digestive health including colorectal cancer, gastrointestinal inflammation, and aids probiotics and other adverse health conditions [13]. Though, some evidence suggests that the effect of bioactive compounds on the human body is moderate and may last over relatively short periods but they could contribute significantly if consumed routinely as part of the daily diet [14]. Therefore,to facilitate discussion on this issue, this review focuses on the therapeutic role of seaweed-derived compounds as a nutraceutical or functional food ingredients for health maintenance and diseases prevention. It examines the existing scientific knowledge on primary and secondary metabolites and functional properties for health-related conditions, their bioavailability and action on body metabolism.
2. Bioactive compounds from seaweed
2.1. Polysaccharides
Polysaccharides are described as an energy reserve and structural compounds of all organisms including marine and higher plants. In seaweed, the polysaccharides are the most important macro-molecule which holds more than 80% of its weight. These resistant polysaccharides are termed as dietary fibres, which are not digested in the body, but due to enzymatic action of microorganisms in the gut can ferment to varying degrees [15]. Edible seaweeds contain a remarkably high amount of dietary fibre varying in the range of 23.5% to 64% DW (dry weight) in species like
Codium, Gracilaria, Ulva, Acanthopora, where the level of polysaccharides are higher than wheat bran [16]. Algal polysaccharide differs from terrestrial plant polysaccharides; they comprise of uncommon polyuronides, some are methylated, acetylated, pyruvylated, or sulfated. Among different polysaccharides, sulfated form such as fucoidan, fucan sulfate, ulvan and carrageenan are most investigated for its biological values [17]. The sulfated polysaccharides (SPS) has wider application in pharmaceutical,nutraceutical and cosmeceutical sectors and they exist in edible seaweeds like ulvan (Chlorophyta), fucoidan (Phaeophyta) and carrageenan (Rhodophyta). The SPS compound exhibit antioxidant, anticancer, anti-inflammatory, antidiabetic, anticoagulant,immunomodulatory and anti-HIV activities [10,11]. These activities are extensively attributed to the interaction between polysaccharide and intestinal microorganisms to prove a functional and medicinal property of sulfated polysaccharides [18]. For example,cytokine modulation, apoptosis inhibition, and protein tyrosine phosphatase inhibition are common examples of SPS and intestinal microorganisms. Furthermore, antioxidant activity and free radical scavenging activity of these polysaccharides are extensively studied against hydroxyl radical (OH?), nitric oxide radical (NO?),superoxide radical (O2??), DPPH, FRAP and ABTS. A recent study has shown SPS is more potent nitric oxide scavenger compared to synthetic antioxidants like BHT and BHA [17,23]. The mechanism of action for this property is derived from their structural feature of sulfate compound and type of sugar moiety attached,molecular weight, and glycosidic linkage. Few researchers reported that the molecular weight of SPS would determine the activity from high to low molecular weight, which in turn reduce the antioxidant activity [19]. In most cases, lower molecular weight possesses more antioxidant activity compared to high molecular weight because proton donor action takes place in the cells are effective in low molecular weight SPS [18,19]. Moreover, this antioxidant property is important to prevent the free radical formation in the cell thereby inhibiting oxidative cell membrane damage.
2.2. Protein and amino acids
The proportion of protein in seaweed ranges up to 45% DW and it differs from species, season and geographical area. Protein content is found to be high in red algae (20%–35%) followed by green (10%–20%) and brown algae (<10%). But some species of green algae like Ulva reticulate, Ulva lactuca, Ulva fasciata and Enteromorpha were reported with high protein content (12%–23%)compared to other species [20]. However, the protein concentration of the seaweed is comparatively higher than terrestrial plant source like soybean, pulses etc [21]. The essential amino acids are present with some free amino acid fractions in the range of 6.47%–24%, especially in A. nodosum (34.4 g/100 g protein),F. vesiculosus (25.1 g/100 g protein) and Ulva spp. (27.0 g/100 g protein). Frequently recorded amino acids are glutamic acid,taurine, threonine, arginine, alanine, and aspartic acid; however,some limiting amino acids such as phenylalanine and methionine were not detected in some seaweeds like C. crispus, Gracilaria sp., O. pinnatifida, Porphyra spp. [22]. The ratio of essential amino acid and non-essential amino acid (EAA/NEAA) is higher in red seaweed (0.98–1.02) followed by green seaweed (0.72-0.97) and brown seaweed (0.73) [20]. However, Viera et al. [23]recorded extraordinarily higher values of EAA/NEAA score from A. nodosum (1.00–1.06) and Gracilaria spp. (1.47–1.74) in the literature so far. Therefore, edible seaweeds are considered as a replacement for an animal protein with a wide range of essential amino acids and the values are similar to legumes and soybean,30%–40% DW [24]. As a result, this level is comparable to RDI(recommended dietary intake) requirements of dietary protein for adults. Other protein derivatives like lectins from Bryothamnion triquetrum and phycobiliproteins (phycoerythrin, phycocyanin and allophycocyanin) from P. yezoensis have gained recent research focus [25]. These proteins have shown host-pathogen interactions against bacteria, virus, and immunomodulation reactions which induces apoptosis in anticancer activity [25].
2.3. Lipids and fatty acids
Seaweed lipids mostly contain long-chain fatty acids, especially polyunsaturated fatty acids (PUFA) with 18- and 22- carbon atoms, depending on species. In general, lipid in seaweed ranges from 0.4% to 5% DW, with abundant saturated fatty acids (SFA)and palmitic acid in all species. Likewise, essential fatty acids(EFA) and PUFA were found abundant in brown seaweeds followed by green and red seaweeds. PUFA greatly varied in seaweed species and the ratio of omega-6-fatty acids and omega-3-fatty acids are of specific interest in the healthy metabolic profile of cardiovascular and obesity-related issues. Omega 3 fatty acids are found to be abundant in Ulva, Acanthophora and Gracilaria species[20], which is extensively reported for anti-inflammatory, antihypertensive, antihyperlipidemic and inhibition of angiotensin I-converting (ACE-I) enzyme [25]. Furthermore, Ulva species contains eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA),the precursor of α-linolenic acid (ALA; 18:3) derived through elongation and desaturation, which articulated to the functional property of anti-inflammatory and antioxidant activity [26]. However, this ratio depends on the amount of nitrogen loaded in the cultivation site of seaweed. The higher amount of nitrogen on the site had an attractive combination of n-6/n-3 (0.3) and 18:2n-6/18:3n-3 (0.5) proportions and a substantial amount of 18:4n-3, 20:5n-3 and 22:5n-3 unsaturated fats mainly reported in Ulva species. These lipids derived from neutral lipid fraction which consists of 62% and the rest was occupied with phospholipids(38%) by high 16:0 (49.6%), 18:1n-7 (14.6%) and 18:3n-3 (13.6%)unsaturated fats [26]. Therefore, Ulva species could be futuristic food ingredients with the perfect combination for health applications.
2.4. Vitamins
Vitamins are the most essential micronutrients to trigger many metabolic pathways and serve as a precursor for enzyme co-factors.Since higher organisms do not have a metabolic pathway to synthesize these cofactors, thus must receive from an external source.Algae are the richest source of vitamins almost contain all essential and non-essential vitamins in it. Numerous seaweeds like Porphyra umbilicalis, Himanthalia elongata and Gracilaria changii contains a high level of vitamin C compared to land vegetables. For instance,seaweed Eisenia arborea contains 34.4 mg/100 g DW vitamin C contents relatively higher than mandarin oranges [27]. Vitamin B12 is also synthesized by prokaryotes which are closely associated with a eukaryotic algal surface that further enhance the B12 level in seaweed. The Vitamin B12 content reported in microalgae Chlorella and Spirulina (33.3 and 15.3 μg/kg FW (fresh weight)) is much higher than the macroalgae nori (1 μg/kg FW). Although found to be low levels in nori, almost 60% of active vitamin B12 coenzymes aggregated would cover the daily need of biologically active vitamin B12 if it is fortified in a smoothie [28]. This result shows that algal food is an alternative source of vitamin B12 for the vegetarian population as most of the higher plants cannot synthesize B12 but it is prevalent in seaweed. A study conducted by Takenaka et al. [29]found that nori fed to vitamin B12 deficient rats had yielded 1.9-fold increase hepatic B12 levels compared to non-seaweed diet. Commercial processing methods such as air drying and lyophilization could alter vitamin uptake in the system, for example, air-dried Pyropia tenera has produced biologically inactive B12, whereas lyophilization process might have improved nutritional quality[28]. Vitamin A (carotenoids) found in seaweeds such as Codium fragile and Gracilaria chilensis is relatively higher than in carrots. The total carotenoids of seaweed range between 25–100 μg/g DW, like P. pavonica consist of a high level of carotenoids whereas C. lentillifera, K. striatum and E. denticulatum were in the range of 33–65 μg/g DW [23]. Beta-carotene and lycopene are carotene derivatives while canthaxanthin, zeaxanthin, violaxanthin, siphonaxanthin and astaxanthin are xanthophyll derivatives[30]. A seaweed-derived lutein and zeaxanthin have been reported for protection against macular degeneration. These dietary pigments are profoundly valuable nutraceutical ingredient which shows unique metabolic activity in human health [24]. They possess powerful antioxidant activities and are closely associated with anticancerous, antihypercholesterolemic and neurodegenerative diseases [8]. According to Ganesan et al. [31], siphonaxanthin from green seaweed is an effective inducer of apoptosis in HL-60 cells compared to fucoxanthin. Fucoxanthin ranging from 1.3 mg/g to 2.4 mg/g DW was predominantly found in brown seaweeds such as wakame, laminaria and sargassum species, while zeaxanthin was major carotenoid compound reported in G. asiatica and G.livida (80.2 μg/g). All these seaweeds are edible in nature and are a major dietary source of carotenoids with high biological values [31].
2.5. Phytochemicals
Seaweeds possess a wide range of secondary metabolites and is continuously involved in the interest of the scientific community for their potential bioactivity, compared to land plants. Generally,the most studied phytochemicals in seaweeds are phlorotannins, gallic acid, quercetin, phloroglucinol, carotenoids and its derivatives. Polyphenol compounds were frequently reported in all genera of seaweeds but most likely their presence is found high in brown and red seaweeds [32]. Most important phenolic compounds derived from marine plants are phlorotannins which assembled through polymerization of phloroglucinol units (1, 3,5-trihydroxybenzene monomer unit) to form polyphloroglucinols[7]. Brown algae possess a high amount of phloroglucinol and used as a nutraceutical agent for health benefits. These phenolic compound reported in H. elongata, Euchema cottonii, Sargassum muticum and Ecklonia Cava, showed potential biological activities like antioxidant, antiproliferative, anti-inflammatory, antidiabetic,anti-HIV, anti-Alzheimer’s activity in vitro [11,33]. Phlorotannins are reported as effective against free radical scavenging effect and metal chelators, this characteristic feature helps to inhibit lipid peroxidation [34]. According to Li et al [34], antioxidant property of phlorotannins are more efficient compared to α-tocopherol in a linoleic model system in vitro, this shows high activity against DPPH, superoxide, peroxy radical assay.
3. Bioavailability of compounds
3.1. Bioavailability of seaweed microelements
The bioaccessibility of micronutrients is influenced by numerous elements such as micro-constituent and types of processing done on the seaweed. Chlorophyll is considered one of the important nutrients in animal metabolism [2]. Three chlorophyll-rich seaweeds were tested for its stability during in vitro digestion which shows that bioavailability of seaweed chlorophyll was found high in nori (?8%) followed by Kombu and sea lettuce [35]. However,the bioavailability of land plants like spinach puree and peas was 5%–10% and 2%–5% respectively, this outcome shows that seaweed chlorophyll is equivalent to the land plants. Nevertheless, the heavy metal accumulation found to be high in marine plants in contrast to the land plants. Seaweeds are known for the high accumulation of heavy metals, as their polysaccharides had anionic carboxyl,sulfate and phosphate linkages that are found to be an appropriate site for heavy metals retention [36]. The bioaccessibility of heavy metals from C. racemosa seaweed collected from 3 different sites of Spain was tested in vitro, there was high bioaccessibility recorded for manganese (Mn) in the range between 71.8%–85.3%,copper (Cu)- 44.3%–56.3%, zinc (Zn)- 37.7%–47.4% and cadmium(Cd)- 41.8%–46.7%. Although, this seaweed shows very low bioaccessibility for lead (Pb) and iron (Fe) which was 22.3%–32.0% and 11.5%–16.5%, respectively [37]. Two forms of Laminaria digitata obtained from the European Market were tested for bioaccessibility which displayed Zn in the range of 77- 31%, Cu- 43% and Mn- 22%–75%, but bioaccessibility of Fe was not determined due to its traceable amount in digestion; thus the results obtained from C. racemosa found better as compared to L. digitata. Further,the cooked seaweed Kombu, Wakame, Sea Lettuce and Nori are reported to contain less amount of metals like Fe- 22.9–556 μg/g,Cu- 1.01–11.7 μg/g and Zn- 7.14–35.8 μg/g. This demonstrated that metals are discharged into the cooking water during heat treatment with the exception of Ni and Zn in Kombu, Cr, Fe and Co inWakame, and Zn in Nori [38]. The fresh warm water soaking treatment at 32°C decreased the iodine content in S. latissima below the threshold value of 2000 mg/kg and treatment of A. esculenta in hypersaline at 2.0 mol/L NaCl concentration decreased the relative cadmium (Cd) content up to 0.5 mg/kg DW [39]. These two types of treatment influenced the nutrient content of the biomass,represented by significant varieties in DW and the substance of bioactive mixes e.g. minerals, polyphenols and fucoxanthin. Therefore, soaking in fresh warm water brought about 49% loss of DW alongside the relative increment of Cd and carbohydrates which binds with an intercellular matrix such as alginate, glucose and fucose (reflecting laminarin and fucoidan levels) in A. esculenta[39]. In vitro digestibility study on cooked seaweed sample (sea spaghetti and Gelidium) shows high dialyzability of iodine (17%)[37]. All these studies showed that enormous amount of elemental compounds found in the varying degree of availability as compared to RDI, but the bioavailability and bioaccessibility depend on body physiological function and composition of diet consumed in human samples. For example, supplementation of Ulva reticulata incorporated chocolate was given to anaemic adolescent girls showed expansion in haemoglobin, MCH (mean cell haemoglobin)values (27.59 mg) might be ascribed to high iron per cent in U. retic-ulata (56 mg/100 g), which displays an appreciable amount of iron bioavailability in the gastrointestinal system [40].

Table 1 Some of the active compounds and their chemical structures found in seaweed.
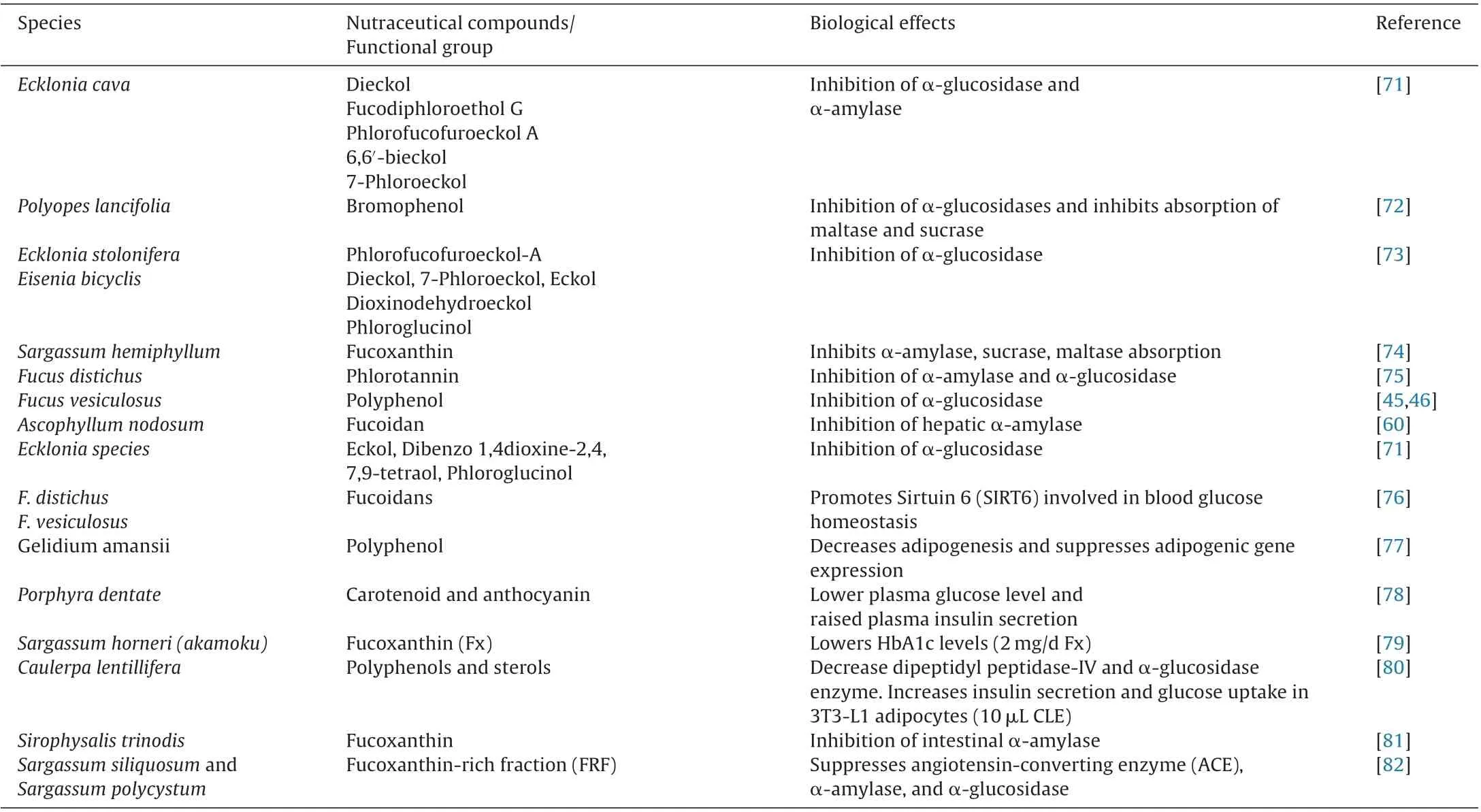
Table 2 Effect of seaweed on glucose metabolism.
3.2. Bioavailability of macro elements
Seaweeds are frequently reported for macromolecules and bioactive compounds and numerous researches have been carried out on isolation and structural elucidation of these compounds.Table 1 summarized some of the active compounds and their chemical structures reported in seaweed. Green seaweed Ulva lactuca and Enteromorphoa contains 20% DW of protein and low yield of polyphenols this would interact with hydrogen bonding of proteinphenol [41]. The high amount of polyphenol might inhibit protein absorption since many brown seaweeds, some red and green seaweed possesses a high amount of phenolic compounds such as phlorotannins, catechol, quercetin that may lead to low bioavailability of protein in vivo. In addition, the protein concentration is often estimated using nitrogen to protein conversion factor, this conversion factor calculated in crude biomass of algae was even lower average factors of 4.59 (red algae), 5.13 (green algae) and 5.38 (brown algae) [22]. However, the biological value of seaweed protein is not clearly articulated during their absorption in the gut.With regard to fat absorption, seaweed possesses 0.2%–5% of lipidenriched with DHA, EPA and omega-3-FA [41]. Although the lipid amount constitutes lower, this will not be underrated since this exhibits nutraceutical property for promoting health. Porphyra tenera showed more than 30 volatile compounds mainly consists of trans-beta-ionone, hexadecanoic acid and 2,6-nonadienal of 20.9%,9.2% and 8.7%, respectively, which inhibits lipid peroxidation in the system [42]. According to Arterburn [42], algal oil DHA is equivalent to salmon DHA and its bioavailability ratio is almost the same. DHA thus represent a safe for human consumption and a convenient source for the vegetarian population.
4. Therapeutic properties of seaweed-derived compounds
4.1. Effect on glucose metabolism
Many new drugs have evolved for diabetes in recent years including oral hypoglycemic agent as well as insulin mimickers. Bioactive compounds from seaweed are found safe and effective against type-2-diabetes, which reverse carbohydrate metabolism enzymes [43]. Among the various seaweed bioactive,alkaloids, flavonoids, carotenoids, polyphenols and phlorotannins were shown to have a hypoglycemic effect (Table 2). Maeda et al.[44] reported that fucoxanthin significantly promotes insulin sensitiveness and reduces blood glucose level in diabetic mice. Fucoidan from brown algae Fucus vesiculosus and Ascophyllum nodosum were frequently reported for reduction of blood glucose level in an animal model [45]. This research demonstrated that low molecular weight of fucoidan trigger the production of beta cells, and limits the secretion of glucagon from alpha-cell in the reverse of blood glucose. Similarly, fucoidan from A. nodosum has shown significant action on α-amylose activity as compared to its counterpart F. vesiculosus fucoidan; this outcome indicates the purity of fucoidan from A. nodosum species had a main role thus showed potential biological activity [46]. Further, α-glucosidase activity relatively depends on species-specific and harvesting period. For instance, fucoidan at low concentration of 0.025 mg/mL extracted from A. nodosum during autumn season showed 80% α-glucosidase inhibition in contrast with fucoidan obtained in other seasons[45]. Similarly, other researchers also identified comparable result on α-glucosidase activity, as fucoidan extracted from A. nodosum showed progressively intense inhibitor of α-glucosidase, with IC50of 0.013 to 0.047 mg/mL than fucoidan from F. vesiculosus(IC50= 0.049 mg/mL). Additionally, fucoidan from F. vesiculosus did not restrain α-amylase action, while fucoidan from A. nodosum diminished α-amylase by 7%–100% at 5 mg/mL concentration level[46]. The mechanism of action of fucoidan molecules is unknown but few interpretations can be made for its inhibitory action which includes (1) strong electrostatic interaction between sulfate groups with negative charge and enzyme, (2) high viscosity of fucoidan may impact on diffusivity in a solvent which might lead to increase in time to reach the enzyme. Additionally, the biological action of seaweed fucoidan also depends on its molecular cutoff, i.e. low molecular weight fucoidan (LMF) has a higher degree of absorption and bioavailability than high molecular fucoidan (HMF) [47].Furthermore, crude seaweed extract also showed intense action on glucose metabolism i.e methanolic extract of E. stolonifera(MEE) strongly inhibits α-glucosidase. It has been proven that MEE significantly reduced plasma glucose and lipid peroxidation in genetically modified non-insulin dependent diabetic KK-Ay male mice. Further, analysis of MEE extract in HPLC-DAD and LC/MS showed that phlorotannin phenolic compound was responsible for the hypoglycemic effect [48]. Further, some research proved that polyphenolic compounds are more potent hypoglycemic than polysaccharide fractions of seaweeds. The polyphenolic fraction of A. nodosum shows the highest α-glucosidase inhibition at low concentration but polysaccharide fraction fails to decrease this level at the same concentration [46].

Fig.1. Proposed mechanism for inhibition of the proliferation of melanoma cells by FCSPs: Activation of macrophages via membrane receptors, which leads to the production of cytokines that enhance NK cell activation. Activated NK cells release Granzyme B and perforin through granule exocytosis into the space between NK cells and melanoma cells to initiate caspase cascades in melanoma cells. Assimilation of Granzyme B by the tumour cells is facilitated by perforin. Granzyme B then initiates apoptosis by triggering the release of mitochondrial cytochrome c and apoptosome formation leading to caspase-3 activation, which in turn translocates the nucleus causing DNA fragmentation—the distinct morphological change of cells by apoptosis. Adopted from [56].
4.2. Effect on cell proliferation
Cell proliferation is the process wherein the number of cell increases due to cell division and cell growth which is the common occurrence in tumours or cancer. Evidence suggested that fucoidan can act as an anti-proliferative agent by inducing the maturation of dendritic cells, in association with other cytokines and modulate the human immune system [12]. Fig.1 represents the proposed mechanism for inhibition of the proliferation of melanoma cells by fucoidans. Macrophages are activated via membrane receptors namely TLR-4, CD14, CR-3 and SR which leads to the production of cytokines such as IL-12, and IFN-γ that enhances NK cell activation which stimulates T-cell activation [49]. Some studies have reported that inhibition of cancer cell depends on the sulfate group of fucose,sulfate content of fucoidan and MW of fucoidan which influences the anti-proliferative effect [5]. Fucoidan obtained from seaweed such as Saccharina cichorioides, Fucus evanescens and Undaria pinnatifida significantly suppress the proliferation of human colon cancer cells (DLD-1) and display less cytotoxicity towards normal mouse epidermal cells (JB6 C141) [50]. However, Ermakova et al.[50] had proved that there was no correlation between fucoidan and sulfate linkage, as fucoidan isolated from E. cava, S. hornery and C. costata with various proportions of sulfate and without sulfate linkage were showed similar cell proliferation against human colon cancer cells (SK-MEL-28), melanoma (DLD-1) [51]. Furthermore, anticancer activity has been relatively proved in secondary metabolites of seaweeds such as phlorotannins, flavonoid, catechol,carotenoids, quercetin and myricetin. Polyphenol-rich Eucheuma cottonii (ECME) was tested against breast cancer cell and after inoculation of 72 h, it was observed that ECME was more active against estrogen-dependent cells than estrogen-independent cells. This outcome showed the presence of polyphenol in ECME was more effective against cancer-associated receptors and the mechanism of gene expression in cancer cell death [51]. Epidemiological studies also indicated that consumption of seaweed lowers the incidence of ovarian, breast and endometrial cancer in the Japanese population compared to the other parts of the world [24].
4.3. Effect on adipose tissue
Obesity is defined as the unwanted accumulation of fat in the body and white adipose tissue (WAT), which inhibits the secretion of cytokine in adipose tissue and leads to a chain of other disorders like diabetes, hypercholesterolemia and stroke [52]. Thermogenesis plays a crucial role in the regulation of obesity mechanism. Some thermogenic protein like uncoupling protein (UCP1, UCP2, and UCP3) families associated with oxidative phosphorylation in brown adipose tissue (BAT). Further, UCP1 deficiency in mice reported being increases resistant to obesity [53]. However, in human, the amount of BAT present in the body is very limited as compared to WAT, which consists of fat accumulation (Fig.2). Therefore, the expression of UCP1 on WAT attains more importance in recent research for anti-obesity therapies. Brown seaweed U. pinnatifida reduced WAT in Wistar rat and KK-Ay mice after 4 weeks of treatment (2% Undaria lipid-fed rats), it was interesting to know that significant reduction in body weight with no variation in food intake of rats [54]. Similarly, U. pinnatifida extract was given to C57BL/6 J mice reduces plasma leptin level and epididymal adipose tissue [55]. This extract consists of fucoxanthin remarkably reduced adipocyte size, fasting blood glucose and insulin level as compared to obese rats [56]. Other species like L. japonica and L. ochotensis consists of fucoxanthin inhibits the absorption of fat and serum triglyceride level in vivo model and also proved an anti-obesity effect on mice [57].
4.4. Effect on lipid metabolism
Epidemiological research confirms a strong association between non-starchy polysaccharide (dietary fibre) and lipid metabolism,the diet rich in these polysaccharides would improve the health status of the colon [58]. The wholesome seaweed wakame in long-term administration (1% in diet) enacts the peroxisome proliferator-activated receptor pathway pursued by β-oxidation,and gluconeogenesis [59,66]. This property was reported in secondary metabolites of seaweed such as ulvan, carrageenan,alginate, fucoidan and fucoxanthin [60]. Ulvan (U) from Ulva pertusa with varying molecular weight (U1- 151.6 Da and U2-28.2 kDa) was tested against hypercholesterolemic rat models showed anti-hypercholesterolemic action with a significant reduction in cholesterol level in an animal model. This shows ulvan has the ability to bind on lipid molecules which reduces total cholesterol and LDL cholesterol (45.2% and 54.1%) significantly. Both U1 and U2 shows elevated serum HDL cholesterol by 22% to 61%as compared to control cereal diet [58]. Thus, MW of seaweed polysaccharides also plays a role in the modification of lipoprotein proportion in lipids. High MW polysaccharides react with LDL cholesterol whereas low molecular weight acts on triacylglycerol and HDL cholesterol. Austin et al. [59] proved that water extract of A. nodosum contains both polysaccharide and polyphenol while the ethanol extract contains rich polyphenol fraction only but later inhibits lipase more efficiently to prevent cholesterol absorption. While Chater et al. [60] reported that concentrates from F. vesiculosus were considerably more intense lipase inhibitors than A. nodosum and Pelvetia canaliculata extracts. In addition, carrageenan has the ability to impersonate texture and sensory quality of lipid, thus it minimizes the total fat absorption in food [61].The main cause of this property is due to the occurrence of nonproteinaceous amino acid-like taurine in red algae (i.e 1.0–1.3 g taurine/100 g) [62]. Although taurine is a non-essential amino acid, consumption of sulfated polysaccharide increases excretion of faecal bile acids and lower cholesterol in the bloodstream [63].Further, the inclusion of seaweed meals to experimental animals increases faecal fat and decreases fat digestibility in animal models.Nori and Wakame seaweeds are shown to improve fermentation (i.e. short-chain fatty acids) in the colon which inhibits lipid emulsification [64]. Similarly, nori and Kombu diet had lower postprandial lipidemia (3 h) levels in albino rats; further Kombu diet showed relatively higher lipidemia values than nori fed counterparts [65]. This shows that metabolization of chylomicron found to be high in seaweed diet with the fact that nori has more solubility than Kombu which induces hypocholesterolemic effects in rats as compared to cellulose fed rats. This cause of action is due to improvement in arylesterase activity which involves lipoprotein metabolism and inhibits lipo-peroxidation in LDL [66]. This tendency is common in hypercholesterolemia where increased peroxidation and hepatic uptake of HDL in the reverse cholesterol transport pathway. The overall report shows that seaweed triggers key reaction in lipid metabolism which includes (1) modifying emulsification of fat in bile acid, (2) disruption of micelle formation, (3) alteration in lipase enzyme, (4) binding on cholesterol site and (5) improve fermentation of colonic bacteria. Apart from polysaccharides, other bioactives such as polyphenol, fucoxanthin and polyunsaturated fatty acids from various seaweed species also influenced the lipid metabolism through a different mode of action(Table 3).
5. Conclusion and future prospects

Fig.2. Effects of fucoxanthin on thermogenesis and lipolysis: the muscle (a) and the adipose tissue (b). Fucoxanthin plays an anti-obesity effect mainly by stimulating uncoupling protein-1 (UCP-1) expression in white adipose tissue (WAT). This protein, situated in the mitochondrial inner cellular membrane, is usually found in brown adipose tissue (BAT) and it is not expressed in WAT in absence of any stimulation. Physiologic bodily metabolism determines heat production: this process is named thermogenesis and UCP-1 dissipates the pH-gradient generated by oxidative phosphorylation, releasing chemical energy as heat. Fucoxanthin was found to promote not only UCP1 protein and mRNA expression but also β3-adrenergic receptor (Adrb3), which is responsible for lipolysis and thermogenesis. This increased sensitivity to sympathetic nerve stimulation may lead to a further up-regulation of fat oxidation in WAT. This adaptive thermogenesis plays a crucial role in energy expenditure as heat, in order to limit weight gain and to favour weight loss. Adopted from [51].

Table 3 Effect of seaweed on lipid metabolism.
Seaweeds have been ascribed as potential sources of functional compounds derived from secondary metabolisms such as fucoxanthin, phlorotannin, fucoidans, laminarin, carrageenan, alginate and agar. These compounds are widely used in food applications for various properties to improve the quality of food. Though seaweeds have been used as a functional ingredient in a commercial application as a stabilizer, emulsifier, thickening agent,texture modifier and phytochemicals enriched with vitamins and dietary fibre, substantial effort is required to establish their role and application in health-promoting foods for direct consumption.Comprehensive knowledge of therapeutic food products as well as the emerging trend towards the development of the functional food industry, seaweed products could have enormous potential as they are linked with high vitamin, high fibre, high minerals and high omega-3-fatty acids which in turn could provide high commercial scope. Several in vitro studies have proved the efficacy of fortified food products from seaweed bioactive compounds however the most challenging factor in the food industry is to develop new products which could appeal the consumers where unfamiliar products are approaching them. However, more in-vivo research is required to support the claims that seaweed could be an alternative source of health supplements which are helpful for the prevention of diseases and disorders. Additionally, it is equally important to enhance public awareness and bring seaweed into the spotlight for developing a more comprehensive and readily available alternative source of ingredients for daily food with therapeutic effects. A number of studies have been carried out to evaluate the food, pharmaceutical and nutraceuticals properties, but more research is needed to understand the safety, toxicity and environmental impact of seaweed cultivation, processing and bioactive mining for commercial scale-up. Additionally, seaweed ingredients based functional food and nutraceutical products should be tested for the presence of contaminants, allergens, heavy metals or hazardous substances generated during seaweed cultivation or processing. These products should comply and adhere to strict safety legislation in order to push forward with their commercial development and production. Future work should also be focused on sustainable processing of seaweed biomass (with the aim to improve resource efficiency) by designing cleaner pre-treatment techniques for enhancing the productivity, industrial scalability and profitability as well as generating zero waste. Therefore, a systematic biorefinery approach to recover a variety of commercial compounds from seaweed biomass with no leftover should be developed/explored.
Some edible seaweed frequently reported in this review paper are only the commonly explored ones for nutraceutical purposes,while the sea is an unexploited pool of nutrient materials for medicinal food in various ways. Despite the great effort in developing new products, not many industrial competent products have been developed utilizing these nutraceuticals for health and wellbeing of human. Primarily this is because extensive human trials and nutritional intervention studies are pre-requisites to establish seaweed as an excellent raw material for nutraceuticals at commercial level. Further studies need to be designed to explore the bio-accessibility and bioavailability of these bioactive compounds for long-term beneficial effects. In addition, cost-effective cultivation technologies, upstream processing of the biomass, greener and environmental friendly extraction approaches/technologies are required to recover these active molecules from seaweed biomass.
Funding
This work was supported by Science Foundation Ireland (SFI)[grant number: 14/IA/2548].
Declaration of Competing Interest
The authors declare that they have no competing interests.
Acknowledgement
None.

- 食品科學(xué)與人類健康(英文)的其它文章
- Almond cold-pressed oil by-product as ingredient for cookies with potential health benefits: Chemical and sensory evaluation
- Impact of hot alkali modification conditions on secondary structure of peanut protein and embedding rate of curcumin
- Mixed vitamin C and zinc diet supplements co-administered with artemether drug improved haematological profile and survival of mice infected with Plasmodium berghei
- Fermentation-enabled wellness foods: A fresh perspective
- Hyperinsulinemia, cancer and maqui berry: The promise of nutritional supplementation
- Antioxidant peptides encrypted in flaxseed proteome: An in silico assessment