
Identification of lineariifolianoid A as a novel dual NFAT1 and MDM2 inhibitor for human cancer therapy
2016-06-05 08:55:01JiangJiangQinSushantaSarkarSukeshVorugantiRajeshAgarwalWeiWangRuiwenZhang
Jiang-Jiang Qin, Sushanta Sarkar, Sukesh Voruganti, Rajesh Agarwal, Wei Wang, Ruiwen Zhang,?
1Department of Pharmaceutical Sciences;2Cancer Biology Center, School of Pharmacy, Texas Tech University Health Sciences Center, Amarillo, TX 79106, USA;3Department of Pharmaceutical Sciences, Skaggs School of Pharmacy and Pharmaceutical Sciences;4University of Colorado Cancer Center, University of Colorado Denver, Aurora, CO 80045, USA.
Identification of lineariifolianoid A as a novel dual NFAT1 and MDM2 inhibitor for human cancer therapy
Jiang-Jiang Qin1,2, Sushanta Sarkar1, Sukesh Voruganti1, Rajesh Agarwal3,4, Wei Wang1,2, Ruiwen Zhang1,2,?
1Department of Pharmaceutical Sciences;2Cancer Biology Center, School of Pharmacy, Texas Tech University Health Sciences Center, Amarillo, TX 79106, USA;3Department of Pharmaceutical Sciences, Skaggs School of Pharmacy and Pharmaceutical Sciences;4University of Colorado Cancer Center, University of Colorado Denver, Aurora, CO 80045, USA.
There is an increasing interest in development of novel anticancer agents that target oncogenes. We have recently discovered that nuclear factor of activated T cells 1 (NFAT1) is a novel regulator of the Mouse Double Minute 2 (MDM2) oncogene and the NFAT1-MDM2 pathway has been implicated in human cancer development and progression, justifying that targeting the NFAT1-MDM2 pathway could be a novel strategy for discovery and development of novel cancer therapeutics. The present study was designed to examine the anticancer activity and underlying mechanisms of action of lineariifolianoid A (LinA), a novel natural product inhibitor of the NFAT1-MDM2 pathway. The cytotoxicity of LinA was first tested in various human cancer cell lines in comparison with normal cell lines. The results showed that the breast cancer cells were highly sensitive to LinA treatment. We next demonstrated the effects of LinA on cell proliferation, colony formation, cell cycle progression, and apoptosis in breast cancer MCF7 and MDA-MB-231 cells, in dose-dependent and p53-independent manners. LinA also inhibited the migration and invasion of these cancer cells. Our mechanistic studies further indicated that its anticancer activities were attributed to its inhibitory effects on the NFAT1-MDM2 pathway and modulatory effects on the expression of key proteins involved in cell cycle progression, apoptosis, and DNA damage. In summary, LinA is a novel NFAT1-MDM2 inhibitor and may be developed as a preventive and therapeutic agent against human cancer.
lineariifolianoid A, NFAT1, MDM2, breast cancer, p53, apoptosis
Introduction
Recent advances in research on the etiology and pathogenesis of human cancers have demonstrated that a number of oncogenes and tumor suppressor genes play a pivotal role in cancer onset, development, progression, and response to therapy[1-6], including the Mouse Double Minute 2 (MDM2) oncogene andTP53tumor suppressor[5-6]. A multitude of targeted therapy approaches have been developed to inhibit the oncogenes[7-9]. The amplification and overexpression of MDM2 are common genetic events in human cancers and correlated with tumor initiation, growth, metastasis, and responses to chemotherapy and radiationtherapy[10-12]. MDM2 plays a major role in cancer cell survival and proliferation, cell cycle progression, apoptosis, migration, and invasion by regulating a variety of signaling pathways[5-6,13-16]. We and others have demonstrated that MDM2 exerts its oncogenic activityviap53-dependent and p53-independent mechanisms, inspiring the development of several different MDM2 targeting approaches[17-24]. Not surprisingly, there is an increasing interest in developing specific, potent, and safe MDM2 inhibitors for cancer prevention and therapy.
There are several MDM2 inhibitors being developed to inhibit the MDM2-p53 interaction, protecting p53 from MDM2-mediated ubiquitination and proteasomal degradation[17-19,23,25-26]. However, concerns have been raised about the limitations on both the efficacy of these MDM2 inhibitors in p53-mutant cancers and the potential toxicity associated with p53 activation[5-6,14]. In continuous efforts to develop novel MDM2 targeting approaches and the specific and potent MDM2 inhibitors, we and others have identified antisense oligonucleotides and siRNAs[27-33], natural products[22,34-39], and synthetic compounds[20-21,40-41], in order to directly inhibit MDM2 expression and/or its activity, emphasizing the p53-independent mechanism. More recently, we have identified that the transcription factor nuclear factor of activated T cells 1 (NFAT1) activates MDM2 expression in a p53-independent manner[42]. NFAT1 has been demonstrated to play an important role in cancer development and progression and has been suggested as a promising molecular target for cancer treatment and prevention[4,43-45].
We have recently proposed to develop MDM2 inhibitorsviatargeting the NFAT1-MDM2 signaling pathway[4,39,42], leading to the identification of lineariifolianoid A (LinA) (Fig. 1) through the screening of our in-house compound library (Data not shown). LinA is a naturally occurring sesquiterpenoid dimer from the aerial parts ofInula lineariifoliaTurcz., which has been used in traditional Chinese medicine "Jinfeicao" for treating digestive disorders, bronchitis, inflammation, and diabetes[46-48]. In our previous study, LinA was discovered as a specific and potent MDM2 inhibitor, but the precise mechanisms by which LinA inhibits MDM2 remain unclear[22]. The present study was designed to evaluate thein vitrocytotoxicity of LinA in a variety of cancer cell lines and the underlying mechanism(s) of action. Since breast cancer cells were highly sensitive to the treatment of this compound, our major efforts were devoted to further investigate its anticancer activity in breast cancer cells. It was hoped that our results from the present study would provide a basis for further development of LinA for human breast cancer therapy in preclinical and clinical settings.
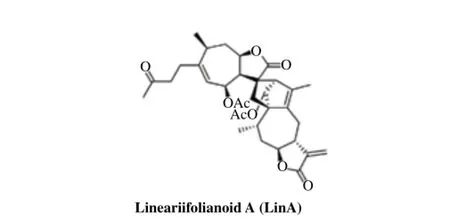
Fig. 1The chemical structure of LinA.
Materials and methods
Chemicals, antibodies, and reagents
LinA was prepared as described in our previous study[48]. All chemicals and solvents used in the present study were of highest grade available. Fetal bovine serum (FBS) was obtained from Atlanta Biologicals (Lawrenceville, GA, USA). The penicillin/streptomycin preparation was bought from Corning (Manassas, VA, USA). The anti-human NFAT1 (1/NFAT-1) antibody was purchased from BD Biosciences (San Jose, CA, USA). The antibodies against human MDM2 (Ab-2), p21 (Ab-1), and p-H2AX (Ser139) were sourced from EMD Chemicals (Gibbstown, NJ, USA). The antibodies against human p53 (DO-1), Cdk2 (M2), Cdk4 (H-22), Cdk6 (C-21), cyclin D1 (DCS-6), cyclin E (HE12), c-Myc (0.N.222), Bax (N-20), Bcl-2 (100), PARP (H-250), Chk1 (G-4), Chk2 (B-4), and ATR (N-19) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The antibodies against human p-Chk1 (Ser317), p-Chk2 (Thr68), and p-p53 (Ser15) were purchased from Cell Signaling Technology (Danvers, MA, USA). The anti-human β-actin (AC-15) antibody was bought from Sigma (St. Louis, MO, USA) and the goat anti-mouse IgG (H+L) and goat anti-rabbit IgG (H+L) antibodies were obtained from Bio-Rad (Hercules, CA, USA).
Cells and cell culture
Human normal and cancer cell lines used in the present study were purchased from American Type Culture Collection (Rockville, MD, USA). All cell culture media were supplemented with 10% FBS and 1% penicillin/streptomycin, except for those indicated otherwise. Human breast cancer cell lines tested in the present study included MCF7, MCF7 p53 knockdown (MCF7p53-/-), MDA-MB-468, and MDA-MB-231. The MCF7 and MDA-MB-231 cells were cultured in Dulbecco's modified Eagle's medium (DMEM). The MCF7p53-/-cells were developed and maintained as reported previously[38,49]. The MDA-MB-468 cells were grown in DMEM/Ham's F12 media. Human prostate cancer cell lines included PC3, LNCaP, and DU145. PC3 cells werecultured in Ham's F12 medium. LNCaP and DU145 cells were grown in Roswell Park Memorial Institute (RPMI) 1640 medium. Human lung cancer A549 cells were cultured in Ham's F12 and H385 cells were grown in RPMI 1640. Human pancreatic cancer cell lines included HPAC, Panc-1, and BxPC3. HPAC cells were grown in DMEM/Ham's F12. Panc-1 and BxPC3 cells were cultured in RPMI 1640. Human colon cancer HCT116 and HCT116 p53 knockdown (HCT116p53-/-) cells were grown in McCoy's 5A. Human osteosarcoma U2OS cells were grown in DMEM. Human ovarian cancer SKOV-3 cells were maintained in McCoy's 5A and OVCAR-3 cells were grown in RPMI 1640 supplemented with 20% FBS, 10 mmol/L of sodium pyruvate, 10 mmol/L of HEPES, 10 mg/L of bovine insulin, and 4.5 g/L of glucose. Human liver cancer HepG2 and Hep3B cells were grown in DMEM. Human glioma U87MG cells were grown in Eagle's minimum essential medium. Human embryonic kidney HEK293 cells were cultured in DMEM. Human breast epithelial MCF10A cells were grown in DMEM/Ham's F12 containing 5% horse serum, 20 ng/mL of EGF, 0.5 mg/ mL of hydrocortisone, 100 ng/mL of cholera toxin, 10 μg/mL of insulin, and 1% penicillin/streptomycin. The inducible MDM2 overexpression (OE) and NFAT1 OE MCF7 cell lines were established and maintained as reported previously[38,42]. In brief, both cell lines were grown in DMEM containing 10 μg/mL of blasticidin and 200 μg/mL of zeocin (Invitrogen, Grand Island, NY, USA). Both MDM2 and HA-NFAT1 plasmids were transfected into cells using the same protocols as we reported previously[35,42].
Cell viability assay
The cytotoxicity of LinA was assessed as described in our previous studies[46,50]. Briefly, the cells were seeded into 96-well plates (3-4×103cells/well) for 24 h and then treated with various concentrations of LinA (0-50 μmol/L) for additional 72 hours. After exposure of cells to LinA, 10 μL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solution (5 mg/mL; Sigma) was added into each well for a 3-hour incubation at 37oC. Absorption was measured at 570 nm using a SYNERGY Mx microplate reader (BioTek, Winooski, VT, USA). The cell survival percentages were calculated by comparisons with the DMSO treated cells.
Cell proliferation assay
The effects of LinA on cell proliferation were evaluated as described previously[21,49]. In brief, cells were grown in 96-well plates at a density of 5-8 × 103cells per well for 24 hours and then exposed to various concentrations of LinA (0, 5, 10, and 25 μmol/L) for another 24 hours. Bromodeoxyuridine (BrdU) was incubated with the cells for 10 hours before the termination of the experiments. Absorption was recorded at the dual wavelengths of 450/540 nm using the SYNERGY Mx microplate reader. The cell proliferation index was calculated by comparing with the DMSO treated cells.
Colony formation assay
The effects of LinA on colony formation were evaluated as reported earlier[21,51]. Briefly, the cells were seeded into 6-well plates (1000 cells/well) and exposed to LinA (5 or 10 μmol/L) for 24 hours. After the removal of the medium containing the tested compound, the cells were cultured in fresh medium for another 10 days. The colonies were fixed, stained, and counted as reported previously[21,51].
Cell cycle analysis
The effects of LinA on cell cycle distribution were analyzed as described previously[20,49]. In brief, the cells were seeded into 6-well plates (2-3 × 105cells/well) and treated with various concentrations of LinA (0, 5, and 10 μmol/L) for 24 hours. The cells were harvested and fixed in physiological saline (0.9% NaCl) and 95% ethanol at 4oC overnight. The cells were then incubated with RNase (Sigma), stained with propidium iodide (Sigma), and analyzed by flow cytometry.
Cell apoptosis assay
The effects of LinA on cell apoptosis were evaluated as described in our previous studies[46,52]. In brief, the cells (2-3 × 105cells/well) were exposed to various concentrations of LinA (0, 5, 10, and 25 μmol/L) for 48 hours. Then, the cells were collected, suspended in Annexin V binding buffer, and incubated with Annexin V-FITC and propidium iodide in the dark at room temperature for 30 minutes. Both early apoptosis and late apoptosis were determined by flow cytometry.
Wound healing assay
The effects of LinA on cell migration were analyzed as reported previously[20,38]. In brief, the monolayer of MDA-MB-231 cells was grown to confluence in 6-well plates and scratched in each well by using a pipette tip. The cells were then exposed to LinA (2 or 5 μmol/L), and five fields of each wound were monitored and photographed at 0, 8, and 24 hours.
Transwell invasion assay
The effects of LinA on cell invasion were examined as described in our previous study[53]. In brief,MDA-MB-231 cells was serum starved overnight and then seeded onto the upper well of Boyden chamber (1-2×104cells/well). After incubation for 6 hours, the cells were exposed to LinA (2 or 5 μmol/L) for another 24 hours. The cells on the upper surface were then removed by wiping with a cotton swab and the cells adhering to the filter were stained with Mayer's Hematoxylin and Eosin solution and photographed under a phasecontrast Olympus microscope (Olympus America Inc). The positive staining area was measured using image analysis software.
Western blotting analysis
In brief, the cells were seeded into 6-cm dishes at 3-4 × 105cells per dish for a 24-hour incubation and treated with various concentrations of LinA (0, 5, 10, and 25 μmol/L) for another 24 hours. Then, the cells were lysed in NP-40 lysis buffer containing a protease inhibitor mixture (Sigma) and the samples were subjected to Western blotting analysis for examining the expression levels of protein of interest, using the methods described previously[39,53].
Statistical analysis
All the quantitative data were expressed as mean ± SEM from at least three independent experiments. Significance of the differences in comparisons of multiple groups was determined by one-way ANOVA and post-hoc pairwise test when there was only one variable. Two-way ANOVA was used when there were two variables. The differences were considered statistically significant atP≤ 0.05.
Results
LinA exerts remarkable cytotoxicity in various cancer cells with minimal activity in normal cells
Thein vitrocytotoxicity of LinA was tested in 20 cell lines representing nine types of human cancer (breast, prostate, lung, pancreatic, colon, ovarian, and liver cancer, sarcoma, and glioblastoma) and two normal cell lines at different concentrations (0 to 50 μmol/L) for 72 hours. The cell viability and the IC50values are shown inTable 1. Notably, the normal cell lines (HEK293 and MCF10A) were much less sensitive to this compound, indicating a selective cytotoxicity of LinA against cancer cells. The breast cancer cell lines, including MCF7 (p53wild-type), MCF7p53-/-, MDA-MB-231 (p53mutant), and MDA-MB-468 (p53mutant), were among the most sensitive cell lines, with the IC50values ranging from 4.4 to 9.1 μmol/L, indicating that its anti-breast cancer activity was notdependent on p53 status, which is important for discovery of a novel anti-MDM2 inhibitor.
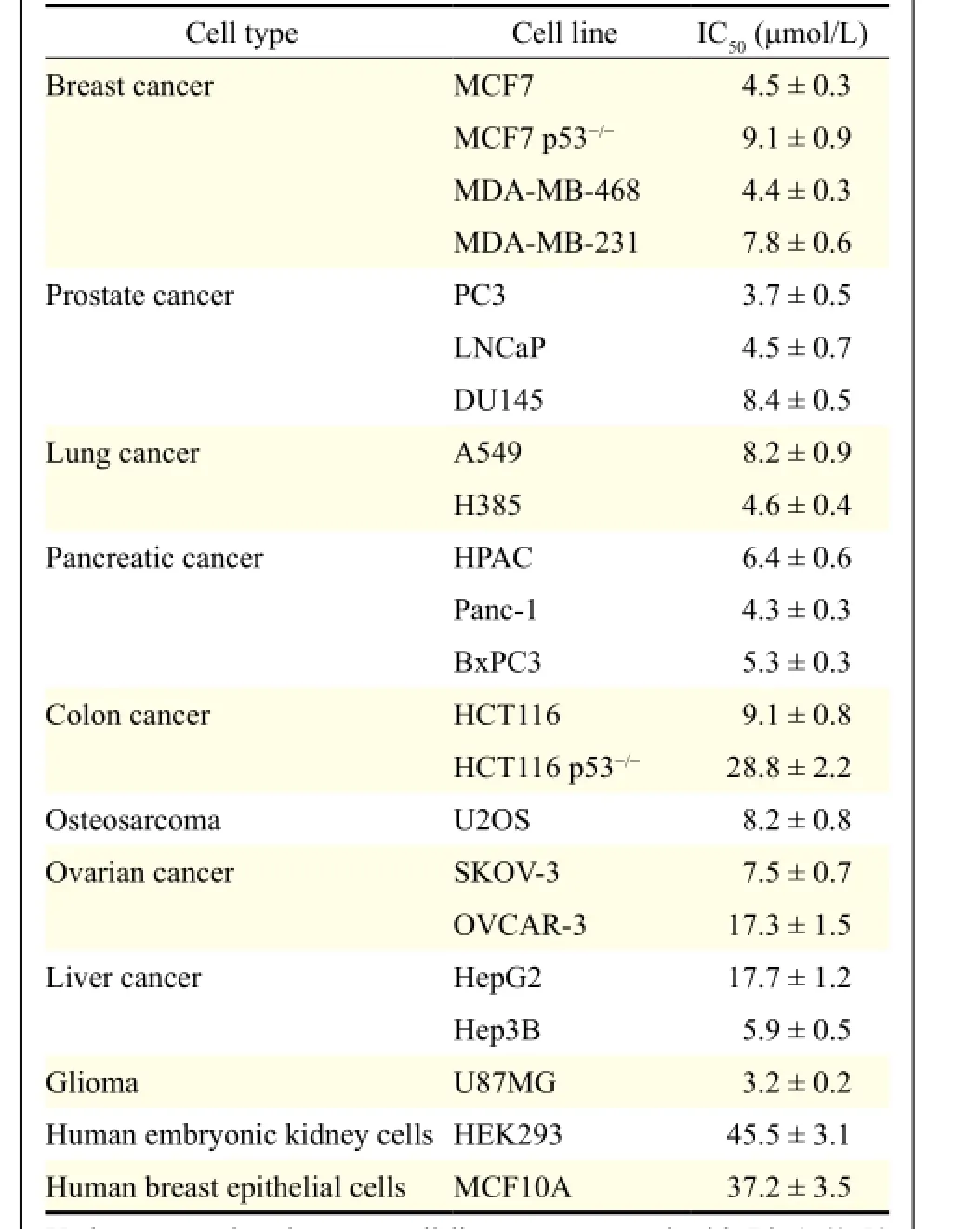
Table 1Cytotoxcity of lineariifolianoid A against normal and cancer cell lines
LinA inhibits cell proliferation and colony formation in breast cancer cells
As shown inFig. 2A, LinA exhibited its antiproliferative effects in both MCF7 and MDA-MB-231 cells in a concentration-dependent manner, regardless of p53 status. At a high concentration of 25 μmol/L, LinA inhibited cell proliferation by 43% (P< 0.01) and 50% (P< 0.01) in MCF7 and MDA-MB-231 cells, respectively. Similarly, LinA also inhibited cell colony formation in both cell lines, in a concentrationdependent manner (Fig. 2B).
LinA delays exit from G2/M phase in breast cancer cells

Fig. 2In vitroanticancer activity of LinA in breast cancer cells. Human breast cancer MCF7 and MDA-MB-231 cells were exposed to various concentrations of LinA for A: 24 hours for the cell proliferation assay, where the proliferation index was calculated by comparing the proliferation of the treated cells with that of DMSO treated cells; B: 24 hours for colony formation assay; C: 24 hours for the cell cycle distribution assay, where the cell cycle distribution was analyzed by comparing the treated cells with that of DMSO trea ted cells; D: 48 hours for cell apoptosis, which was evaluated using the Annexin V-FITC method. All assays were performed in triplicate and repeated three times. (*P< 0.05 and **P< 0.01)
As shown inFig. 2C, LinA delays exit from G2/M phase of the cell cycle in a concentration-dependent manner in both MCF7 and MDA-MB-231 cells, indicating that the effect was independent of p53. LinA at 5 μmol/L markedly increased the number of cells in G2/M phase in both cell lines (P< 0.05). Moreover, in comparison to the control, treatment of cells with 10 μmol/L of LinA led to an accumulation of cells in G2/M phase by 33.1% (P< 0.01) and 48.5% (P< 0.01) in MCF7 and MDA-MB-231 cells, respectively.
LinA induces apoptosis in breast cancer cells
As shown inFig. 2D, LinA induced significant apoptosis in a concentration-dependent manner, while MDA-MB-231 cells were less sensitive than MCF7 cells. After treatment with 25 μmol/L of LinA for 48 hours, 53.2% of MCF7 cells and 14.3% of MDA-MB-231 cells underwent apoptosis, which were significantly higher than that of control cells (bothP< 0.01).
LinA inhibits cell migration and invasion in breast cancer cells
As shown inFig. 3A, untreated MDA-MB-231 cells migrated into the wound area rapidly in a 24-hour treatment period, whereas LinA at the sub-lethal concentrations (2 and 5 μmol/L) significantly inhibited cell migration in a concentration-dependent manner. Similarly, Transwell invasion assay showed that LinA remarkably reduced MDA-MB-231 cell invasion (Fig. 3B). In comparison to the control cells, LinA at 2 and 5 μmol/L reduced the invasion of MDA-MB-231 cells by 41% (P< 0.05) and 87% (P< 0.01), respectively.
LinA inhibits the NFAT1-MDM2 pathway independent of p53
We next explored possible molecular mechanisms of LinA-induced anticancer effects. As shown inFig. 4A, LinA decreased the protein expression of NFAT1 and MDM2 in both MCF7 and MDA-MB-231 cells, in a concentration-dependent manner. LinA also increased the expression level of wild-type p53 in MCF7 cells, but it did not show any effects on the mutant p53 in MDA-MB-231 cells.
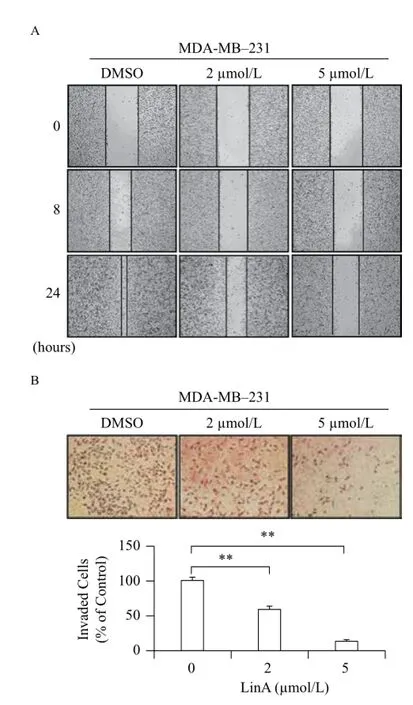
Fig. 3In vitropreventive effects of LinA on breast cancer cell migration and invasion. Human breast cancer MDA-MB-231 cells were exposed to various concentrations of LinA for 24 hours for (A) wound healing assay and (B) Transwell invasion assay. All assays were performed in triplicate and repeated three times. (**P< 0.01)
LinA modulates the expression of key proteins associated with cell cycle progression, apoptosis, and DNA damage response
To further explore the mechanisms of action for anticancer activity of LinA, we next examined the effects of LinA on the expression of key proteins involved in regulating cell cycle progression, apoptosis, and DNA damage response. In both MCF7 and MDA-MB-231 cells, LinA significantly increased the expression level of p21 and decreased the levels of Cdk2, Cdk4, Cdk6, cyclin D1, cyclin E, and c-Myc compared to the control (Fig. 4B). The compound also markedly activated Bax and cleavage of PARP, and decreased Bcl-2 levels in both cell lines (Fig. 4C). The inhibition of ATR by LinA also led to the phosphorylation of Chk1 at Ser317, Chk2 at Thr68, p53 at Ser15, and H2AX at Ser139 (Fig. 4D).
MDM2 overexpression reduces anti-breast cancer activity of LinA
To demonstrate the importance of MDM2 in LinA's anti-breast cancer activity, we compared the effects of LinA on parent and inducible MDM2 overexpression MCF7 cells. As shown inFigs. 5A-5B, Tet treatment resulted in MDM2 overexpression (Fig. 5A), which increased colony formation (Fig. 5B) and attenuated the inhibitory effects of LinA on MDM2 expression (Fig. 5A) and colony formation (Fig. 5B). Similar results were obtained in MDA-MB-231 cells. As shown inFigs. 5C-5F, transient transfection of an MDM2 plasmid caused MDM2 overexpression (Fig. 5C), leading to increased colony formation (Fig. 5D), enhanced cell migration (Fig. 5E) and invasion (Fig. 5F), and mitigated the inhibitory effects of LinA (Figs. 5C-5F) in the cells.
NFAT1 overexpression activates MDM2 and decreases LinA's anticancer activity
To demonstrate the crucial role of the NFAT1 and NFAT1-MDM2 pathway in LinA's anticancer activity, we further compared the inhibitory effects of LinA on parent and NFAT1 OE MCF7 and MDA-MB-231cells. As shown inFigs. 6A-6D, NFAT1 overexpression increased MDM2 expression (Figs. 6A&6C) and colony formation (Fig. 6B&6D) and decreased the inhibitory effects of LinA (Figs. 6A-6D) in both cell lines. In NFAT1 overexpressing MDA-MB-231 cells, NFAT1 overexpression also increased cell migration (Fig. 6E) and invasion (Fig. 6F), and attenuated LinA's inhibitory effects.
Discussion
In the present study, we investigated anticancer activity and the molecular mechanisms of the newly identified NFAT1-MDM2 inhibitor LinA. We have demonstrated several important points that would be of great importance in further research and development of this compound as an anticancer agent. First, LinA displayed potent cytotoxicity against a broad spectrum of human cancer cell lines; breast cancer cell lines were among the most sensitive cell lines, which gave a good explanation for the focus on this cancer type. Second, normal cells were much less sensitive to LinA, suggesting a selective cytotoxicity of this compound. Third, LinA inhibited cell colony formation, delayed exit from G2/M phase of the cell cycle, and induced apoptosis in breast cancer cells in a concentration-dependent manner, regardless of p53 status of the cells. Fourth, thecompound reduced breast cancer cell migration and invasion at sub-lethal concentrationsin vitro. Fifth, LinA inhibited both NFAT1 and MDM2 expression, and modulated the expression of key proteins related to cell cycle progression, apoptosis, and DNA damage response. Finally, enforced MDM2 or NFAT1 expression remained effective when the cells were exposed to LinA.Fig. 7depicts the possible mechanisms of action for LinA.
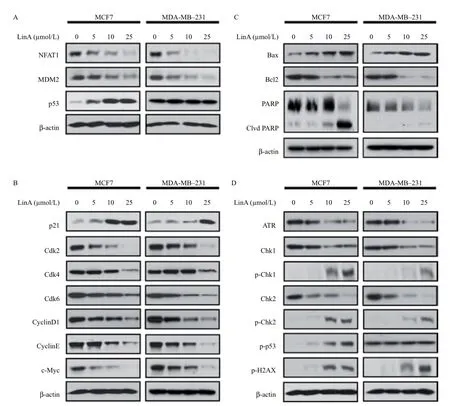
Fig. 4Effects of LinA on the NFAT1-MDM2 pathway and on the expression of key proteins related to cell cycle progression, apoptosis, and DNA damage response in breast cancer cells. Human breast cancer MCF7 and MDA-MB-231 cells were exposed to various concentrations of LinA for 24 hours. Then, the expression levels of (A) NFAT1, MDM2, and p53, and the key proteins related to (B) cell cycle progression, (C) apoptosis, and (D) DNA damage response were examined by Western blotting assay. All assays were performed in triplicate and repeated three times.
Significant advances have recently been made in our understanding of genetic alterations in human cancers, including the amplification, overexpression, and mutation of key oncogenes (e.g.,MDM2andNFAT1) as well as the mutation and deletion of tumor suppressors (e.g.,TP53andBRCA1)[1-2,4-6,54]. Targeted therapies that inhibit the oncogenes and/or reactivate the tumor suppressors offer great promise for patients with advanced cancer[7-9]. The targeted therapies may also enhance chemotherapy-induced cytotoxicity and boost responses to immunotherapy, improving the clinical outcomes of chemotherapy and immunotherapy for patients with advanced diseases[7]. There is an increasing interest in developing novel approaches to inhibit theMDM2oncogene via p53-dependent and p53-independent mechanisms[5-6,13-16]. Considering our recent discovery in the NFAT1-MDMD2 pathway thatactivates MDM2 in a p53-independent manner[4,39,42], we proposed to develop NFAT1-MDM2 inhibitors for cancer therapy, leading to the identification of LinA.
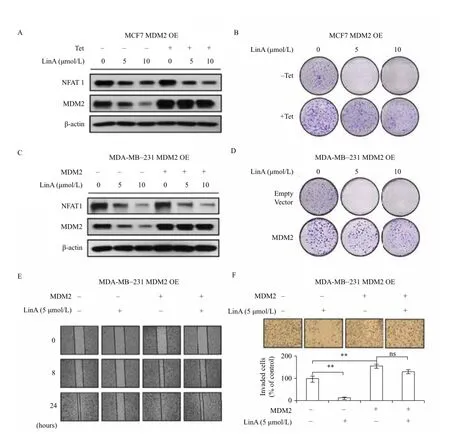
Fig. 5Effects of MDM2 overexpression on LinA's activity. The inducible MDM2 overexpression MCF7 cells were incubated with (+Tet; 1 μg/mL) or without tetracycline (-Tet) for 24 hours and then treated with LinA (0, 5, or 10 μmol/L) for 24 hours for (A) the expression of NFAT1 and MDM2 and (B) the colony formation assay. MDA-MB-231 cells were transfected with an MDM2 plasmid or the empty vector for 24 hours and then treated with LinA (0, 5, or 10 μmol/L) for 24 hours for (C) the expression of NFAT1 and MDM2, (D) the colony formation assay, (E) the wound healing assay (5 μmol/L of LinA treatment), and (F) Transwell invasion assay (5 μmol/L of LinA treatment). All assays were performed in triplicate and repeated three times. (**P< 0.01, "ns" denotes "not significant")

Fig. 6Effects of NFAT1 overexpression on LinA's activity. The inducible NFAT1 overexpression MCF7 cells were incubated with (+Tet; 1 μg/mL) or without tetracycline (-Tet) for 24 hours and then treated with LinA (0, 5, or 10 μmol/L) for 24 hours for (A) the expression of HA-NFAT1 and MDM2 and (B) the colony formation assay. MDA-MB-231 cells were transfected with an HA-NFAT1 plasmid or an HA plasmid (empty vector) for 24 hours and then treated with LinA (0, 5, or 10 μmol/L) for 24 hours for (C) the expression of HA-NFAT1 and MDM2, (D) the colony formation assay, (E) the wound healing assay (5 μmol/L of LinA treatment), and (F) Transwell invasion assay (5 μmol/L of LinA treatment). All assays were performed in triplicate and repeated three times. (**P< 0.01, "ns" denotes "not significant")
In our recent study with cell-based screening assays for identifying novel NFAT1-MDM2 inhibitors, LinA was identified as a novel dual NFAT1 and MDM2 inhibitor from our in-house compound library (Data not shown). We demonstrated potent cytotoxicity of LinA against a wide spectrum of human cancer cell lines, of which breast cancer cell lines exhibited high sensitivity to LinA. These findings are consistent with our previous results that the NFAT1-MDM2 pathway plays a crucial role in breast cancer cell growthin vitroandin vivo[39,42]. We further demonstrated the potent inhibitory effects of Lin A on NFAT1 and MDM2 in both p53-wildtype and -mutant breast cancer cell lines. Given these observations, we then demonstrated the critical role of the NFAT1-MDM2 pathway in LinA's anti-breast cancer activity using MDM2 or NFAT1 overexpressing MCF7 and MDA-MB-231 cells. In the overexpression studies, the enforced MDM2 or NFAT1 expression was established and remained effective when the cells treated with LinA, including decreased colony formation and reduced cell migration and invasion. Interestingly, compared with the parent cells, MDM2 or NFAT1 overexpression reduced the inhibitory effects of LinA on colony formation, cell migration, and cell invasion. Although the exact mechanisms of action are not completely understood, the possible reason may be related to high level of MDM2 or NFAT1, which compete for the limited available compound (LinA) in the assay system. The current overexpression studies may not represent the situation of real MDM2 or NFAT1 overexpression in clinical setting, but may still provide evidence for LinA binding to its targets (MDM2 and NFAT1). Nevertheless, further investigations are required for establishing the link between LinA's activity and its inhibitory effects on the NFAT1-MDM2 pathway.
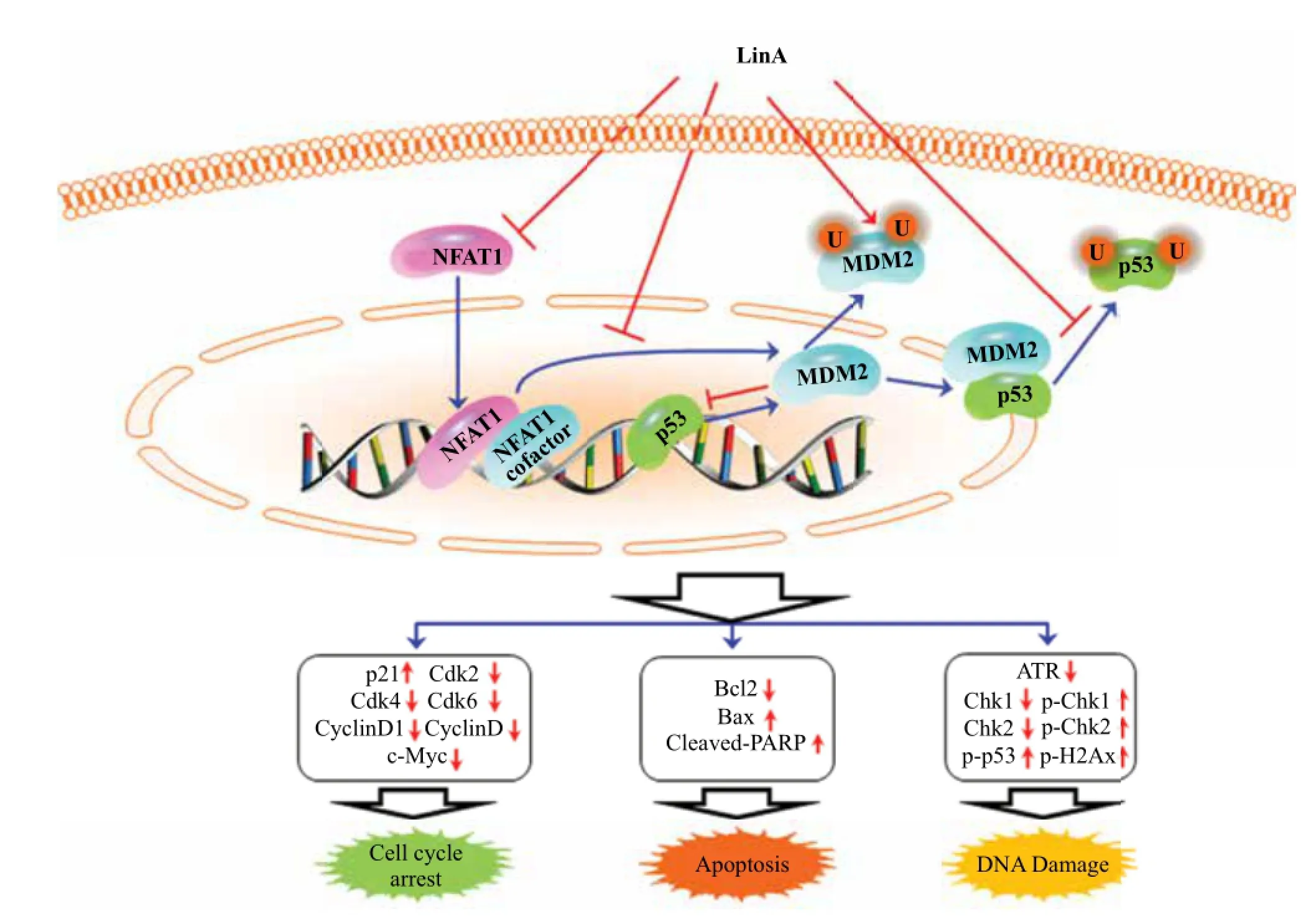
Fig. 7The schematic diagram depicts the possible mechanisms of action of LinA against cancer cells. LinA inhibits cancer cell growth and induces cell cycle arrest and apoptosis by inhibiting the NFAT1-MDM2 pathway and modulating the expression of key proteins involved in regulating cell cycle progression, apoptosis, and DNA damage.
With respect to the therapeutic potency of LinA, our results indicated that this compound exerted its anticancer activity at relatively low concentration, as elucidated in cell viability, colony formation, cell cycle distribution, and apoptosis assays. However, the effects of LinA on cell proliferation were achieved at very high concentration, especially in MDA-MB-231 cells. The dose-response effects could just be a function of cytotoxicity and not a real function in cell proliferation. Depended on the observation, the major mechanisms of cell death might be apoptosis induction in MCF7 cells and cell cycle regulation in MDA-MB-231 cells. In addition, we demonstrated the preventive effects of LinA on cell migration and invasion at sub-lethal concentrations, indicating that LinA might possess therapeutic potential in metastatic cancers. Given the potent anticancer activity, we further examined the effects of LinA on various key proteins involved in regulating cell cycle progression, apoptosis, and DNA damage response. The majority of the proteins have been reported to be associated with the MDM2-p53 and NFAT1-MDM2 pathways. The results are similar to previous observations on other MDM2 inhibitors that were developed to inhibit MDM2 expression. However, the present study just provided the proof-of-conceptin vitrodata for the new compound, but it had certain limitation. In the future, various animal models such as orthotopic mouse models and metastatic models should be employed to evaluate thein vivoefficacy of LinA and characterize its toxicological profile. Additional techniques such as pharmacological and genetic interventions should be utilized to generate more evidence for demonstrating LinA as a specific NFAT1-MDM2 dual inhibitor.
In summary, we have identified LinA, a natural sesquiterpenoid dimer, as a novel NFAT1-MDM2 inhibitor and demonstrated its potent anticancer activity in breast cancer cells. Our results suggest that LinA may be a potential therapeutic agent for the treatment and prevention of human cancers. Future studies should further demonstrate its activityin vivo, when used alone and/or in combination with other conventional cancer chemotherapies. If we could confirm its anticancer effects in preclinical settings, it can be developed as a clinical candidate for breast cancer therapy. It should be pointed out that most advanced breast cancers do not express wild-type p53; our newly identified p53-independent MDM2 inhibitor may offer new hope for the cancer patients without functional p53 expression.
Acknowledgements
This work was supported by the National Institutes of Health (NIH) grant R01 CA186662 (to R.Z.) and CA102514 (to R.A.). The content is solely the responsibility of the authors, and do not necessarily represent the official views of the National Institutes of Health. This work was also supported by American Cancer Society (ACS) grant RSG-15-009-01-CDD (to W.W.).
[1] Qiu B, Simon MC. Oncogenes strike a balance between cellular growth and homeostasis[J].Semin Cell Dev Biol, 2015, 43: 3-10.
[2] McCormick F. KRAS as a therapeutic target[J].Clin Cancer Res, 2015, 21(8): 1797-1801.
[3] Wang W, Qin JJ, Voruganti S, et al. Polycomb group (PcG) proteins and human cancers: Multifaceted functions and therapeutic implications[J].Med Res Rev, 2015, 35(6):1220-1267.
[4] Qin JJ, Nag S, Wang W, et al. NFAT as cancer target:Mission possible[J].Biochim Biophys Acta, 2014, 1846(2):297-311.
[5] Nag S, Qin J, Srivenugopal KS, et al. The MDM2-p53 pathway revisited[J].J Biomed Res, 2013, 27(4): 254-271.
[6] Qin JJ, Nag S, Voruganti S, et al. Natural product MDM2 inhibitors: Anticancer activity and mechanisms of action[J].Curr Med Chem, 2012, 19(33): 5705-5725.
[7] Vanneman M, Dranoff G. Combining immunotherapy and targeted therapies in cancer treatment[J].Nat Rev Cancer, 2012, 12(4): 237-251.
[8] Wong DJ, Ribas A. Targeted therapy for melanoma[J].Cancer Treat Res, 2016, 167: 251-262.
[9] Luo J, Solimini NL, Elledge SJ. Principles of cancer therapy: oncogene and non-oncogene addiction[J].Cell, 2009, 136(5): 823-837.
[10] Santarius T, Shipley J, Brewer D, et al. A census of amplified and overexpressed human cancer genes[J].Nat Rev Cancer, 2010, 10(1): 59-64.
[11] van den Broek AJ, Broeks A, Horlings HM, et al. Association of the germline TP53 R72P and MDM2 SNP309 variants with breast cancer survival in specific breast tumor subgroups[J].Breast Cancer Res Treat, 2011, 130(2): 599-608.
[12] Boersma BJ, Howe TM, Goodman JE, et al. Association of breast cancer outcome with status of p53 and MDM2 SNP309[J].J Natl Cancer Inst, 2006, 98(13):911-919.
[13] Bouska A, Eischen CM. Murine double minute 2: p53-independent roads lead to genome instability or death[J].Trends Biochem Sci, 2009, 34(6): 279-286.
[14] Wade M, Li YC, Wahl GM. MDM2, MDMX and p53 in oncogenesis and cancer therapy[J].Nat Rev Cancer, 2013, 13(2): 83-96.
[15] Kaindl U, Morak M, Portsmouth C, et al. Blocking ETV6/ RUNX1-induced MDM2 overexpression by Nutlin-3 reactivates p53 signaling in childhood leukemia[J].Leukemia, 2014, 28(3): 600-608.
[16] Bohlman S, Manfredi JJ. p53-independent effects of Mdm2[J].Subcell Biochem, 2014, 85: 235-246.
[17] Vassilev LT, Vu BT, Graves B, et al.In vivoactivation of the p53 pathway by small-molecule antagonists of MDM2[J].Science, 2004, 303(5659): 844-848.
[18] Issaeva N, Bozko P, Enge M, et al. Small molecule RITA binds to p53, blocks p53-HDM-2 interaction and activates p53 function in tumors[J].Nat Med, 2004, 10(12):1321-1328.
[19] Shangary S, Qin D, McEachern D, et al. Temporal activation of p53 by a specific MDM2 inhibitor is selectively toxic to tumors and leads to complete tumor growth inhibition[J].Proc Natl Acad Sci U S A, 2008, 105(10):3933-3938.
[20] Wang W, Qin JJ, Voruganti S, et al. The pyrido[b]indole MDM2 inhibitor SP-141 exerts potent therapeutic effects in breast cancer models[J].Nat Commun, 2014, 5: 5086.
[21] Wang W, Qin JJ, Voruganti S, et al. Identification of a new class of MDM2 inhibitor that inhibits growth of orthotopic pancreatic tumors in mice[J].Gastroenterology, 2014, 147(4): 893-902.
[22] Qin JJ, Wang W, Voruganti S, et al. Identification of a new class of natural product MDM2 inhibitor:In vitroandin vivoanti-breast cancer activities and target validation[J].Oncotarget, 2015, 6(5): 2623-2640.
[23] Ding Q, Zhang Z, Liu JJ, et al. Discovery of RG7388, a potent and selective p53-MDM2 inhibitor in clinical development[J].J Med Chem, 2013, 56(14): 5979-5983.
[24] Rew Y, Sun D. Discovery of a small molecule MDM2 inhibitor (AMG 232) for treating cancer[J].J Med Chem, 2014, 57(15): 6332-6341.
[25] Zhang Z, Chu XJ, Liu JJ, et al. Discovery of potent and orally active p53-MDM2 inhibitors RO5353 and RO2468 for potential clinical development[J].ACS Med Chem Lett, 2014, 5(2): 124-127.
[26] Zhang Z, Ding Q, Liu JJ, et al. Discovery of potent and selective spiroindolinone MDM2 inhibitor, RO8994, for cancer therapy[J].Bioorg Med Chem, 2014, 22(15):4001-4009.
[27] Chen L, Agrawal S, Zhou W, et al. Synergistic activation of p53 by inhibition of MDM2 expression and DNA damage[J].Proc Natl Acad Sci U S A, 1998, 95(1): 195-200.
[28] Chen L, Lu W, Agrawal S, et al. Ubiquitous induction of p53 in tumor cells by antisense inhibition of MDM2 expression[J].Mol Med, 1999, 5(1): 21-34.
[29] Wang H, Nan L, Yu D, et al. Antisense anti-MDM2 oligonucleotides as a novel therapeutic approach to human breast cancer:in vitroandin vivoactivities and mechanisms[J].Clin Cancer Res, 2001, 7(11): 3613-3624.
[30] Wang H, Wang S, Nan L, et al. Antisense anti-MDM2 mixed-backbone oligonucleotides enhance therapeutic efficacy of topoisomerase I inhibitor irinotecan in nude mice bearing human cancer xenografts:In vivoactivity and mechanisms[J].Int J Oncol, 2002, 20(4): 745-752.
[31] Wang H, Nan L, Yu D, et al. Anti-tumor efficacy of a novel antisense anti-MDM2 mixed-backbone oligonucleotide in human colon cancer models: p53-dependent and p53-independent mechanisms[J].Mol Med,2002, 8(4): 185-199.
[32] Wang H, Yu D, Agrawal S, et al. Experimental therapy of human prostate cancer by inhibiting MDM2 expression with novel mixed-backbone antisense oligonucleotides:in vitroandin vivoactivities and mechanisms[J].Prostate, 2003, 54(3): 194-205.
[33] Zhang Z, Li M, Wang H, et al. Antisense therapy targeting MDM2 oncogene in prostate cancer: Effects onproliferation, apoptosis, multiple gene expression, and chemotherapy[J].Proc Natl Acad Sci U S A, 2003, 100(20):11636-11641.
[34] Li M, Zhang Z, Hill DL, et al. Genistein, a dietary isoflavone, down-regulates the MDM2 oncogene at both transcriptional and posttranslational levels[J].Cancer Res, 2005, 65(18): 8200-8208.
[35] Li M, Zhang Z, Hill DL, et al. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway[J].Cancer Res, 2007, 67(5): 1988-1996.
[36] Wang W, Wang H, Rayburn ER, et al. 20(S)-25-methoxyldammarane-3beta, 12beta, 20-triol, a novel natural product for prostate cancer therapy: activityin vitroandin vivoand mechanisms of action[J].Br J Cancer, 2008, 98(4): 792-802.
[37] Wang W, Rayburn ER, Zhao Y, et al. Novel ginsenosides 25-OH-PPD and 25-OCH3-PPD as experimental therapy for pancreatic cancer: anticancer activity and mechanisms of action[J].Cancer Lett, 2009, 278(2): 241-248.
[38] Wang W, Zhang X, Qin JJ, et al. Natural product ginsenoside 25-OCH3-PPD inhibits breast cancer growth and metastasis through down-regulating MDM2[J].PLoS One, 2012, 7(7): e41586.
[39] Qin JJ, Wang W, Voruganti S, et al. Inhibiting NFAT1 for breast cancer therapy: New insights into the mechanism of action of MDM2 inhibitor JapA[J].Oncotarget, 2015, 6(32): 33106-33119.
[40] Wang W, Rayburn ER, Velu SE, et al.In vitroandin vivoanticancer activity of novel synthetic makaluvamine analogues[J].Clin Cancer Res, 2009, 15(10): 3511-3518.
[41] Wang W, Rayburn ER, Velu SE, et al. A novel synthetic iminoquinone, BA-TPQ, as an anti-breast cancer agent:in vitroandin vivoactivity and mechanisms of action[J].Breast Cancer Res Treat, 2010, 123(2): 321-331.
[42] Zhang X, Zhang Z, Cheng J, et al. Transcription factor NFAT1 activates the mdm2 oncogene independent of p53[J].J Biol Chem, 2012, 287(36): 30468-30476.
[43] Kaunisto A, Henry WS, Montaser-Kouhsari L, et al. NFAT1 promotes intratumoral neutrophil infiltration by regulating IL8 expression in breast cancer[J].Mol Oncol, 2015, 9(6): 1140-1154.
[44] Quang CT, Leboucher S, Passaro D, et al. The calcineurin/NFAT pathway is activated in diagnostic breast cancer cases and is essential to survival and metastasis of mammary cancer cells[J].Cell Death Dis, 2015, 6: e1658.
[45] Tie X, Han S, Meng L, et al. NFAT1 is highly expressed in, and regulates the invasion of, glioblastoma multiforme cells[J].PLoS One, 2013, 8(6): e66008.
[46] Qin JJ, Jin HZ, Huang Y, et al. Selective cytotoxicity, inhibition of cell cycle progression, and induction of apoptosis in human breast cancer cells by sesquiterpenoids from Inula lineariifolia Turcz[J].Eur J Med Chem, 2013, 68:473-481.
[47] Nie LY, Qin JJ, Huang Y, et al. Sesquiterpenoids from Inula lineariifolia inhibit nitric oxide production[J].J Nat Prod, 2010, 73(6): 1117-1120.
[48] Qin JJ, Huang Y, Wang D, et al. Lineariifolianoids A-D, rare unsymmetrical sesquiterpenoid dimers comprised of xanthane and guaiane framework units from Inula lineariifolia[J].Rsc Advances, 2012, 2(4): 1307-1309.
[49] Yang X, Wang W, Qin JJ, et al. JKA97, a novel benzylidene analog of harmine, exerts anti-cancer effects by inducing G1 arrest, apoptosis, and p53-independent up-regulation of p21[J].PLoS One, 2012, 7(4): e34303.
[50] Voruganti S, Qin JJ, Sarkar S, et al. Oral nano-delivery of anticancer ginsenoside 25-OCH3-PPD, a natural inhibitor of the MDM2 oncogene: Nanoparticle preparation, characterization,in vitroandin vivoanti-prostate cancer activity, and mechanisms of action[J].Oncotarget, 2015, 6(25):21379-21394.
[51] Voruganti S, Xu F, Qin JJ, et al. RYBP predicts survival of patients with non-small cell lung cancer and regulates tumor cell growth and the response to chemotherapy[J].Cancer Lett, 2015, 369(2): 386-395.
[52] Chen D, Wang W, Qin JJ, et al. Identification of the ZAKMKK4-JNK-TGFbeta signaling pathway as a molecular target for novel synthetic iminoquinone anticancer compound BA-TPQ[J].Curr Cancer Drug Targets, 2013, 13(6): 651-660.
[53] Wang W, Cheng J, Qin JJ, et al. RYBP expression is associated with better survival of patients with hepatocellular carcinoma (HCC) and responsiveness to chemotherapy of HCC cellsin vitroandin vivo[J].Oncotarget, 2014, 5(22):11604-11619.
[54] Pennington KP, Walsh T, Lee M, et al. BRCA1, TP53, and CHEK2 germline mutations in uterine serous carcinoma[J].Cancer, 2013, 119(2): 332-338.
?Correspondening author: Dr. Ruiwen Zhang, MD, PhD, DABT, Department of Pharmaceutical Sciences, School of Pharmacy, Texas Tech University Health Sciences Center, 1406 S. Coulter Dr. Suite 1116, Amarillo, TX 79106, USA. Tel: 1-806-414-9248, E-mail: ruiwen.zhang@ttuhsc.edu. ? 2016 by the Journal of Biomedical Research. All rights reserved.
Received 29 January 2016, Revised 22 February 2016, Accepted 03 April 2016, Epub 25 May 2016
R730.53, Document code: A.
The authors reported no conflict of interest.
10.7555/JBR.30.20160018
 THE JOURNAL OF BIOMEDICAL RESEARCH2016年4期
THE JOURNAL OF BIOMEDICAL RESEARCH2016年4期
- THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Internal carotid artery agenesis with stenosed intercavernous anastomosis: a case report
- Fenmented rice bran prevents atopic dermatitis in DNCB-treated NC/Nga rnice
- Pharmacologicai advantages of melatonin in immunosence by improving activity of T lymphocyfes
- Immunogenicity and protective efficacy of DNA vaccine against visceral leishmaniasis in BALB/c mice
- Characerization cfintegrons and novel cassette arrays in bacteria from clinical isloates in china,2000-2014
- Clinical usefulness of ankie brachial indes and brachial-ankie puise wave velocity in patients with ischemic stroke