
DDX3X regulates cell survival and cell cycle during mouse early embryonic development
2014-04-19 00:34:17QianLiPanZhangChaoZhangYingWangRuWanYeYangXuejiangGuoRanHuoMinLinZuominZhouJiahaoSha
Qian Li,Pan Zhang,Chao Zhang,Ying Wang,Ru Wan,Ye Yang,Xuejiang Guo,Ran Huo,Min Lin, Zuomin Zhou,Jiahao Sha
State Key Laboratory of Reproductive Medicine,Nanjing Medical University,Nanjing,Jiangsu 210029,China.
DDX3X regulates cell survival and cell cycle during mouse early embryonic development
Qian Li,Pan Zhang,Chao Zhang,Ying Wang,Ru Wan,Ye Yang,Xuejiang Guo,Ran Huo,Min Lin, Zuomin Zhou,Jiahao Sha?
State Key Laboratory of Reproductive Medicine,Nanjing Medical University,Nanjing,Jiangsu 210029,China.
DDX3X is a highly conserved DEAD–box RNA helicase that participates in RNA transcription,RNA splicing, and mRNA transport,translation,and nucleo–cytoplasmic transport.It is highly expressed in metaphase II(MII) oocytes and is the predominant DDX3 variant in the ovary and embryo.However,whether it is important in mouse early embryo development remains unknown.In this study,we investigated the function of DDX3X in early embryogenesis by cytoplasmic microinjection with its siRNA in zygotes or single blastomeres of 2–cell embryos. Our results showed that knockdown of Ddx3x in zygote cytoplasm led to dramatically diminished blastocyst for–mation,reduced cell numbers,and an increase in the number of apoptotic cells in blastocysts.Meanwhile,there was an accumulation of p53 in RNAi blastocysts.In addition,the ratio of cell cycle arrest during 2–cell to 4–cell transi–tion increased following microinjection of Ddx3x siRNA into single blastomeres of 2–cell embryos compared with control.These results suggest that Ddx3x is an essential gene associated with cell survival and cell cycle control in mouse early embryos,and thus plays key roles in normal embryo development.
DDX3X,early embryo,p53,apoptosis,cell cycle
INTRODUCTION
In mammals,when an ovulated egg is fertilized,the formation of a new individual occurs.During the maternal–embryo transition,once the zygote starts mitosis and divides into a 2–cell embryo,the embryo transitions from maternal gene to zygotic gene depen–dence.The degradation of maternal mRNAs and proteins is~90%complete at the 2–cell embryo stage and zygotic gene activation(ZGA)starts,which marks zygotic gene transcription;translation then occurs[1,2]. The pre–implantation development of embryo continual cleavage from 1–cell zygote to multi–cell blastocyst comprises the early stage of embryo devel–opment,and is a prerequisite for new individual growth and development.
It is well understood that cell survival maintenance and successful progression of the cell cycle are critical steps for the embryo.As previously reported,many genes and pathways associated with cell survival and cell cycle control play essential roles in early embryo development,including calcium/calmodulin–dependent kinase/CREB transcription factor and AKT(protein kinase B)/MDM2 mediated survival pathways[3],as well as cyclins and cyclin–dependent kinases(cyclin A2,Cdk2[4],and cyclin B[5]).Some persistent maternalfactors and zygotic gene transcripts interact with each other and together contribute to the maintenance of cell survival[6].Transcriptional and translational level con–trol of cell survival,apoptosis,and cell cycle related genes is the chief regulatory mechanism in embryo development;therefore,the subgroups of transcription factors and RNA metabolism regulatory genes are of prime importance during this process.However,the precise genes that play a role remain unclear.
DDX3 is a highly conserved subfamily of the DEAD–box RNA helicase family,and plays important roles in RNA metabolism,including RNA transcrip–tion,RNA splicing,mRNA transport,translation initiation,and cell cycle regulation[7].In mice,there are three members of the DDX3 subfamily that exhibit high similarity:DDX3X,DDX3Y,and PL10[8];Ddx3x in mouse exhibits 99%similarity to DDX3X in humans.DDX3X is ubiquitously expressed in various tissues,and it is the predominant DDX3 variant in the ovary and embryo[9],indicating that it may play an indispensible role in RNA metabolism in the ovary and embryo.DDX3 has been reported to function in cooperation with translation initiation factors including eIF4A,eIF4E,eIF4F and PABP1 and to play a role in protein synthesis in yeast[10]and mammalians[11–13];it may control the transcriptional or translational levels of key cell cycle regulators(including cyclin B,cyclin A,cyclin D1 and chk1)[14–16],thus controlling the cell cycle.It participates in CRM1–dependent nucleo–cyto–plasmic export pathway[17,18]and regulates many pro–teins that exhibit important functions in development. It has also been reported that the orthologues of DDX3 in urochordates and several invertebrates are highly expressed in germ cells and newblasts and are indispensible in asexual reproduction and embryogen–esis[19,20].
In our previous study on mouse oocyte proteomics, we demonstrated the presence of both non–phosphory–lated and phosphorylated forms of DDX3X in mouse oocytes using two dimension(2–DE)electrophoresis and fluorescently labeled Pro–Q Diamond dye[21]. Because the proteins in metaphase II(MII)oocytes are known to be important for subsequent early embryo development,we hypothesized that DDX3X may play a role in the process.Moreover,DDX3X is a highly conserved protein,and is indispensible in embryogen–esis in low organisms;thus,further investigation con–cerning its potentially important role and function in mouse early embryo development is warranted.
In this study,we found that expression of DDX3X was decreased after oocyte fertilization,and increased again following 4–cell stage embryo.We investigated the function of Ddx3x in mouse early embryo by zygote cytoplasmic microinjection of Ddx3x small interfering RNA(siRNA)and observed subsequent embryo development in vitro to blastocyst stage.Our findings showed that after Ddx3x knockdown,blasto–cyst formation was markedly hindered,the number of cells was reduced,and apoptosis signals were increased in blastocyst embryos;additionally,there was an accumulation of p53 in the RNAi groups com–pared with the control.Our data suggest that DDX3X is a key regulator in pre–implantation embryo development; the function is potentially induced via the p53 pathway, which directly induces apoptosis and cell cycle arrest.
MATERIALS AND METHODS
Mice
ICR strain mice were obtained from SLAC Laboratory Animal Co.,Ltd.(Shanghai,China),and maintained in an environment of 22°C,12/12 hours light/dark cycle,and 50–70%humidity.Animal care and experimental procedures were conducted accord–ing to the Animal Research Committee guidelines of Nanjing Medical University.The study protocol was approved by the local institutional review board at the authors′affiliated institution.
Collection of oocyte,zygote,and culture of early embryo
Collecting MII oocytes,6–8–week–old female ICR mice were superovulated by intraperitoneal injection with 10 IU pregnant mare serum gonadotropin (PMSG)followed by 10 IU human chorionic gonado–tropin(hCG).Approximately 16 hours after hCG injection,mice were sacrificed and cumulus–enclosed MII oocytes were obtained by tearing the ampulla of oviducts in HEPES–buffered CZB medium.Cumulus cells were removed following brief exposure to 300 IU/mL of hyaluronidase(Sigma,St.Louis,MO, USA)to denude oocytes.
To obtain zygote and 2–cell embryo,superovulated female ICR mice were mated with ICR males just after hCG injection.About 24 or 44 hours after hCG injection,zygotes or 2–cell embryos were harvested in HEPES–CZB medium from oviducts,respectively. Thoroughly washed early embryos were cultured in CZB medium under mineral oil(Sigma)at 37°C in a 5%CO2atmosphere for subsequent development.
Immunofluorescence and confocal microscopy
Embryos were fixed in 4%paraformaldehyde in PBS for 30 minutes at room temperature and then per–meabilized in PBS containing 0.5%Triton X–100 for30 minutes at 37°C.After two washes with PBS con–taining 0.1%Tween 20 and 0.01%Triton X–100, embryos were incubated in blocking solution(1% BSA in PBS)for 1 hour and then hybridized with 1:300 DDX3X antibody(rabbit polyclonal antibody, ab62581,Abcam,Cambridge,MA,USA)diluted in blocking solution for 1 hour.After three washes, embryos were labeled with 1:100 FITC(Fluorescein isothiocyanate)conjugated goat anti–rabbit secondary antibody(Beijing Zhongshan Biotechnology Co., China)for 45 minutes and then counterstained with propidium iodide or Hoechst for 5 minutes.Finally, embryos were mounted onto slides with 1,4–diazabi–cyclo[2.2.2]octane(DABCO)and examined using a Zeiss 510 laser confocal microscope(Zeiss Fluorescent Microsystems,Go¨ttingen,Germany).The same parameter settings were used for experimental and control groups.
SiRNA oligoribonucleotides
SiRNA duplex oligoribonucleotides targeting the coding region of Ddx3x(GenBank Accession no: NC_000086)were obtained from Invitrogen(Carlsbad, CA,USA).The siRNA sequences were as follows: #02(sense)5′–GGAAAUACAGAUGCUGGCCCG–UGAU–3′,(antisense)5′–AUCACGGGCCAGCAU–CUGUAUUUCC–3′;#04(sense)5′–GGCA–GAUUCGCUGGAGGAUUUCUUA–3′,(antisense) 5′–UAAGAAAUCCUCCAGCGAAUCUGCC–3′; #09(sense)5′–CCUAGACCUGAACUCUUCA–GAUAAU–3′;(antisense)5′–AUUAUCUGAAGA–GUUCAGGUCUAGG–3′.
Non–silencing siRNA nucleotides were used as a negative control.Zygotes of RNAi and negative con–trol group were cultured in CZB medium for further observation and the developmental status of each group was determined and analyzed using a Nikon TE2000–S microscope(Nikon,Yuko,Japan).
DDX3X siRNA microinjection
SiRNAs were prepared as described above and then microinjected into the cytoplasm of MII oocyte, zygote,or single blastomere of 2–cell embryos as pre–viously described[22],and according to our previous work[23,24].The microinjections were repeated at least three times,and at least 100 oocytes or embryos were used.Nikon Diaphot Eclipse TE 300 inverting micro–scope(Nikon,Yuko,Japan),equipped with Narishige MM0–202N hydraulic three–dimensional micromani–pulators(Narishige Inc.,Tokyo,Japan),was used in these experiments.Approximately 10 pL diluted siRNA was injected into one oocyte or embryo in all experiments.After microinjection,MII oocytes or embryos were washed thoroughly and cultured in CZB medium under mineral oil at 37°C in a 5%CO2atmo–sphere,and observed at specific stages of development.
Real time RT-PCR
Total RNA was extracted from embryos using RNeasy Micro Kit(QIAGEN,Hilden,Germany) according to the manufacturer′s protocol.Real–time polymerase chain reaction(real–time PCR)analysis was conducted using Takara Real Time PCR Kit (Takara Bio,Mountain View,CA,USA)and the β–actin mRNA levels in the same samples were used as an internal control.There were three replicates for each sample for each independent experiment.
Primer sequences were:Ddx3x–forward:5′–CTCCGATTTCTCGGTACTCT–3′;Ddx3x–reverse: 5′–GACTTCCCTCTTGAATCACC–3′;β-actin–for–ward:5′–CCGTAAAGACCTCTATGCC–3′;β-actin–reverse:5′–CTCAGTAACAGTCCGCCTA–3′.
Western blotting assays
Morphologically–normal blastocysts(30 per sample) were lysed with sodium dodecyl sulfate(SDS)sample buffer,and then the proteins were separated by polya–crylamide gel electrophoresis(SDS–PAGE).The immunoblotting procedure was done as routinely per–formed.The protein bands were detected using an Enhanced Chemiluminescence Detection Kit (Amersham,Little Chalfont,UK).Primary antibodies used were:1:1000 of p53(rabbit polyclonal antibody, 10442–1–AP,Proteintech)
Cell counts and TUNEL assay
Morphologically–normal blastocysts of the control and experiment groups were collected and fixed in 4%paraformaldehyde in PBS.Hoechst was used to label the nuclei of embryos.The number of cells per embryo was counted using Nikon TE2000–S fluores–cent microscopy.The counting process was repeated by two individuals who were blind to the results of all other experiments of this study.For the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)assay,blastocysts were collected to detect apoptosis using an In Situ Cell Death Detection Kit (POD;Roche,Nutley,NJ,USA)according to the manufacturer′s protocol.Apoptosis signals were observed with a Zeiss 510 laser confocal microscope.
Statistical analysis

Fig.1The expression of DDX3X in oocytes and different stage early embryos.A:Western blotting assays showed that DDX3X was expressed in the ovary and oocyte;there is a single band around 72 kDa as predicted both in the ovarian protein sample(left panel)and oocyte sample (right panel).B:Immunofluorescence staining of DDX3X in germinal vesicle(GV)oocyte,metaphase II(MII)oocyte and different stages of early embryo from 1–cell zygote to blastocyst.Each image was photographed at the same parameter settings.Scale bar=10 μm.
All experiments were repeated at least three times. The differences between the control and experiment groups were analyzed using the one way ANOVA. Homogeneity of variance test was performed.In the post hoc comparisons,least significant difference (LSD)test was used for comparisons with equal var–iances,and Tamhane′s T2 test was performed for the comparisons with unequal variances.Values of P<0.05 were considered to indicate a statistically sig–nificant result.The data are given as mean±SD.
RESULTS
The expression pattern of DDX3X in mouse ovary and various early embryo stages
In our previous study on mouse oocyte proteomics, we demonstrated the presence of DDX3X in mouse oocytes using two–dimension electrophoresis(2–DE)[21].In the present study,DDX3X was confirmed to be expressed in mouse ovary and MII oocyte by Western blot analysis using total mouse ovarian protein or oocyte protein samples.As shown inFig.1A,there was a single band approximately 72 kDa as predicted in both the ovary and oocyte.We then further explored the expression pattern of DDX3X in GV(germinal vesicle stage)oocyte,MII oocyte,and different stage early embryos by immunofluorescence staining.We found that DDX3X was expressed in the cytoplasm and nucleus in GV oocyte,and the intensity was increased in MII oocyte,but largely declined after fer–tilization in 1–cell zygote and 2–cell embryo;however, in subsequent stages,there was an intense fluorescent DDX3X signal in both the cytoplasm and nucleus from the 4–cell stage to the blastocyst stage(Fig.1B).
Knockdown ofDdx3xby zygote cytoplasmic microinjection of siRNA markedly diminished blastocyst formation
As Ddx3x exhibited increasing and higher expres–sion levels in early embryo stages,we hypothesized that it may play a role in early embryogenesis. Knockdown of a target gene by microinjection of its siRNA in the cytoplasm of zygote followed by obser–vation of subsequent embryo development has been successfully used to identify candidate genes that are important during that period of development.Thus, we purchased three pairs of siRNA and initially veri–fied their knockdown efficiency.MII oocytes were injected with Ddx3x siRNA for 18 hours,then oocyte mRNA was extracted immediately using RNeasy Micro Kit and Ddx3x mRNA level was examined by Real Time PCR.As shown inFig.2AandB,the efficiency levels of the three pairs of siRNA were obvious compared with the negative control;among them,#02 and#09 siRNA exhibited a greater knock–down effect compared with#04 siRNA;thus,they were chosen for the subsequent experiments. We further confirmed that the effects of siRNA lasted until the blastocyst stage;there were still marked decreases of Ddx3x in the#02 and#09 siRNA groups compared with control,with an average of 60% and 70%Ddx3x remaining in blastocysts,respectively (Fig.2C).
Zygote cytoplasm microinjection was repeated,and each stage of development was observed and recorded; our findings showed that there was a significant decreaseinblastocystformationrateintheRNAigroups and the negative control group exhibited normal early embryo development compared with the non–treatment group(Fig.3A).As shown inFig.3B,there were minor obstacles when embryos developed to the 4–cell and morula stage,in both the#02 and#09 siRNA groups,but a marked block from the morula to the blas–tocyst stage(41.35%±11.5%and44.6%±11.0%blas–tocyst ratio per total 2–cell embryo in the#02 and#09 siRNA groups,respectively,and 95.4%±4.9%in the negative control group and 99.3%±1.6%in the non–treatment group)(Fig.3B).
Ddx3xknockdown reduced the number cells and an increase in apoptosis in blastocysts
For further investigation of Ddx3x–knockdown blas–tocyst–stage embryos,TUNEL assay and immuno–fluorescence were performed.A large percentage of embryos appeared smaller and abnormal because devel–opmental arrest occurred from morula to blastocyst in the RNAi groups;as shown inFig.4AandC,the apoptosis signals in these embryos were much greater compared with controls(19.5%±11.5%, 17.5%±7.5%,5.6%±3.0%,and 5.5%±3.4%of TUNEL positive cells per total cells in#02,#09, negative control,and non–treatment group,respectively) and there were significant differences between the#02 and#09 RNAi groups compared with the negative control(P<0.001).In addition,cells were counted according to the nucleus number stained by Hoechst, and blastocytsts in the RNAi group contained a fewer number of cells compared with controls(31.0±8.7, 33.3±11.1,59.2±11.9,and 62.8±11.6 cells in#02, #09,negative control,and non–treatment group, respectively).
As knockdown of Ddx3x in zygotes induces apoptosis, we hypothesized that DDX3X plays an important role in early embryos by influencing downstream proteins that control cell survival.As p53 is an marker of apoptosis, we detected the protein level of p53 by Western blot ana–lysis in Ddx3x knockdown and normal blastocysts.The p53 levels were increased in the#02 and#09 RNAi groups in comparison with control(Fig.4D).
Single blastomere injection of 2-cell stage embryo withDdx3xsiRNA led to embryo development arrest
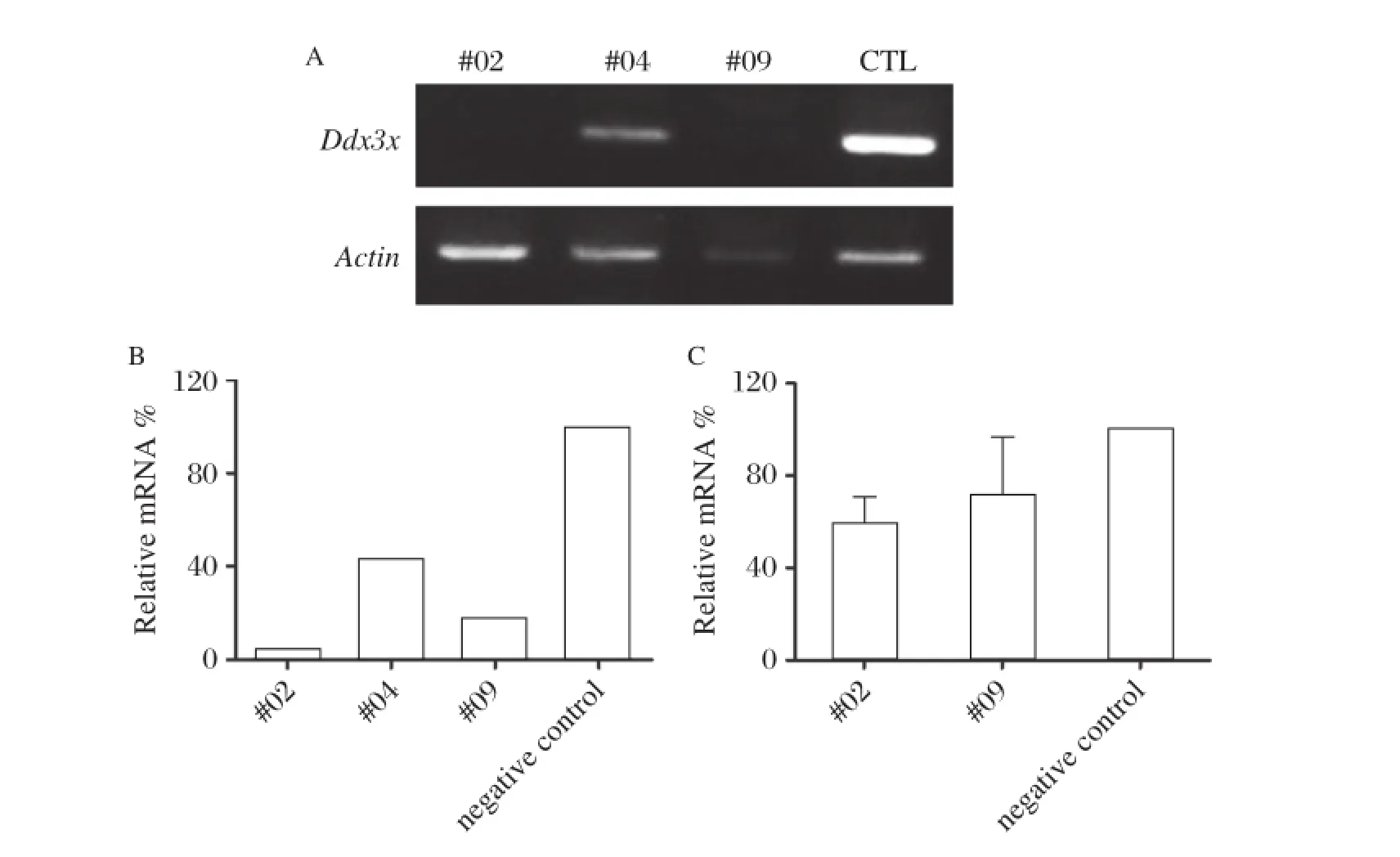
Fig.2Verification of the knockdown efficiency ofDdx3xsiRNA.A:RT–PCR of Ddx3x;the three pairs of siRNA were verified in MII oocytes by microinjection and after 18 hours the RNA of oocytes was extracted and RT–PCR was performed.Non–silencing siRNA was injected as negative control.The knockdown effects were more obvious in#02 siRNA and#09 siRNA compared with#04 siRNA when compared with negative control(CTL).The levels of endogenous β-actin mRNA were used as an internal control.B:Relative expression values for each sample inFig.2Awere normalized to the level of mouse β-actin expression relative to that in negative control.C:The knockdown efficiency of#02 siRNA and#09 siRNA continued until the blastocyst stage after siRNA was injected at the zygote stage.
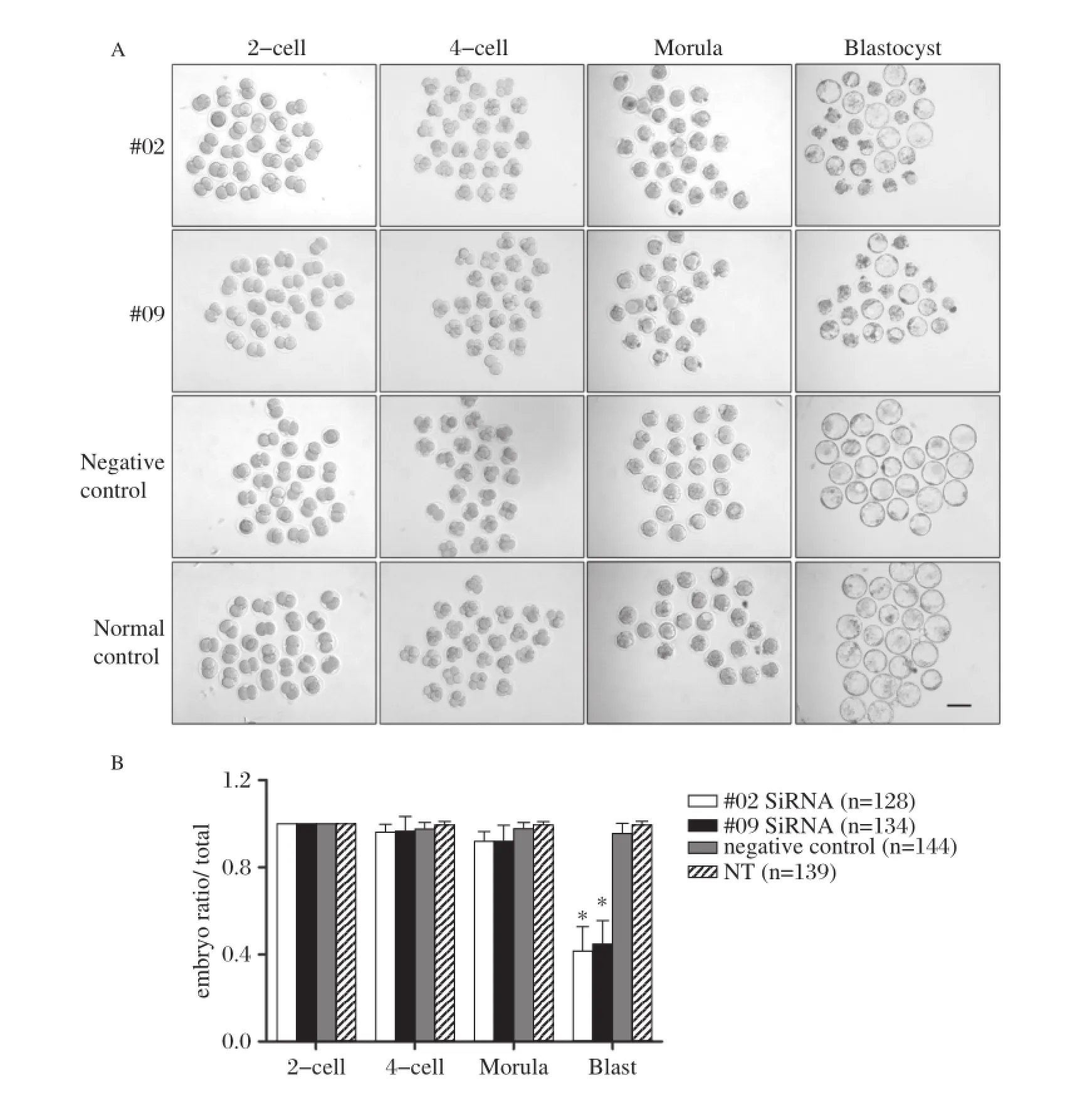
Fig.3 Ddx3xsiRNA microinjection in zygote cytoplasm reduced blastocyst formation.A:Representative images of embryo development in#02 siRNA,#09 siRNA,negative control,and normal non–treatment control.There was impaired blastocyst formation in the#02 siRNA and#09 siRNA groups.B:Repeated experiments were performed and the number of each stage embryo was counted and calculated into four groups(n=128, 134,144,and 139 in#02 siRNA,#09 siRNA,negative control,and non–treatment control,respectively).The blastocyst ratios per total 2–cell embryo of the#02 siRNA and#09 siRNA groups were decreased to 41.4%and 44.6%,respectively.*P<0.001.Scale bar=100 μm.
Ddx3x siRNA with FITC was injected into one of the two blastomeres of 2–cell embryos,and subsequent embryo development was observed using FITC tracing of the knockdown–blastomere.The results showed that there was an increased embryo division arrest ratio from the 2–cell stage to 4–cell stage in the RNAi group compared with the negative control group.As shown inFig.5A,there was a higher ratio of division arrest of FITC–positive blastomeres in all embryos with FITC fluorescence;the data analysis is shown inFig.5B. We further followed the arrested development of 2–cell to 4–cell embryos to the morula stage;in the non–treat–ment group,blastomere division continued to later stages as normally;however,the RNAi blastomeres exhibited arrested development at the single cell stage, and finally apoptosis occurred(Fig.5C).
DISCUSSION
As shown by the immunofluorescence staining of DDX3X in oocytes and various embryo stages, DDX3X is highly expressed in mature MII oocytes, but expression is reduced substantially after fertilization in 1–cell zygotes and 2–cell embryos and then increased again from the 4–cell stage to later stages.We hypothe–size that the decrease in DDX3X is due to maternal fac–tor degradation,particularly in the late 2–cell stage, whereas the increase is heightened by ZGA.
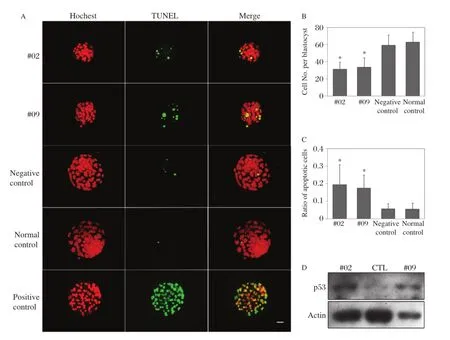
Fig.4 Ddx3xknockdown led to a reduction in the number of cells,an increase in apoptosis signals,and p53 accumulation in blastocysts.A:Immunofluorescence staining and TUNEL assay in the four groups of blastocysts.Red,Hoechst;green,TUNEL signal.B:Cell numbers were counted according to the nuclei stained by Hoechst;blastocysts in the RNAi group contained significantly fewer total cells compared with controls.C:The apoptosis signals were much greater in the RNAi groups compared with controls(n=29,26,29,and 29 in#02 siRNA,#09 siRNA, negative control,and non–treatment control,respectively),***P<0.001.D:Western blot analysis showed an accumulation of p53 in RNAi blastocysts. The same number of blastocysts(n=50)were loaded per panel,and the levels of endogenous β–actin were used as an internal control.Scale bar =20 μm.
We observed that development of morula to blasto–cyst was markedly hampered along with cell number reduction and increase of apoptosis following siRNA microinjection into zygote cytoplasm;additionally, there was an accumulation of p53 in blastocysts. DDX3X is a highly conserved DEAD–box helicase that plays a role in RNA transcription,RNA splicing, translation initiation,CRM1 mediated protein transport, and so on,and there are several potential mechanisms underlying the phenotype:(1)DDX3X has been pre–viously reported to participate in the CRM1 dependent nucleo–cytoplasmic export pathway[12,17,18],which nor–mally transports leucine–rich NES(nuclear export sequences)containing proteins,UsnRNAs,rRNAs and some specific subsets of cellular mRNAs(reviewed by[25–27]).The apoptosis marker p53 protein was reported to be a CRM1–dependent nucleo–cytoplasmic pro–tein[28,29],which is stabilized and retained in the nucleus where it induces the expression of genes involved in cell cycle arrest or apoptosis;it is degraded once transported into the cytoplasm by a CRM1–dependent way.The ability to retain p53 in the nucleus is critical for a cell to produce a p53–mediated stress response[28].As it showed an accumulation of p53 in the Ddx3x RNAi blastocytes,we hypothesize that DDX3X may poten–tially influence the transport of p53 through the CRM1 dependent nucleo–cytoplasmic export pathway and sub–sequently induce the increase of total p53,which finally prevents embryo growth.(2)It is well established that DDX3X participates in transcription and translation of a subgroup of genes that encode cell cycle regulators, including cyclin E1 in HeLa cells[30],cyclin B in yeast[14], cyclin A and cyclin B in hamster cell line ET24[16];once Ddx3x is knocked down,these cell cycle related proteins potentially decrease and lead to cell cycle arrest and finally activate p53–induced apoptosis.(3)DDX3 homologue Ded1p functions in initiating protein synth–esis in yeast,and there is an immediate and markeddecrease of total protein synthesis in ded1 mutants[11,31]. A similar function of DDX3X as a general translation initiation factor has been reported in mammals[32,33]. Although the effects of DDX3 on translation are still controversial,we propose that DDX3X may be also involved intranslationalcontrol inmouse earlyembryos, and finally driving apoptosis in cells in the DDX3X RNAi embryos.However,the ultimate mechanisms of DDX3X function in early mouse embryo development require further research and exploration.
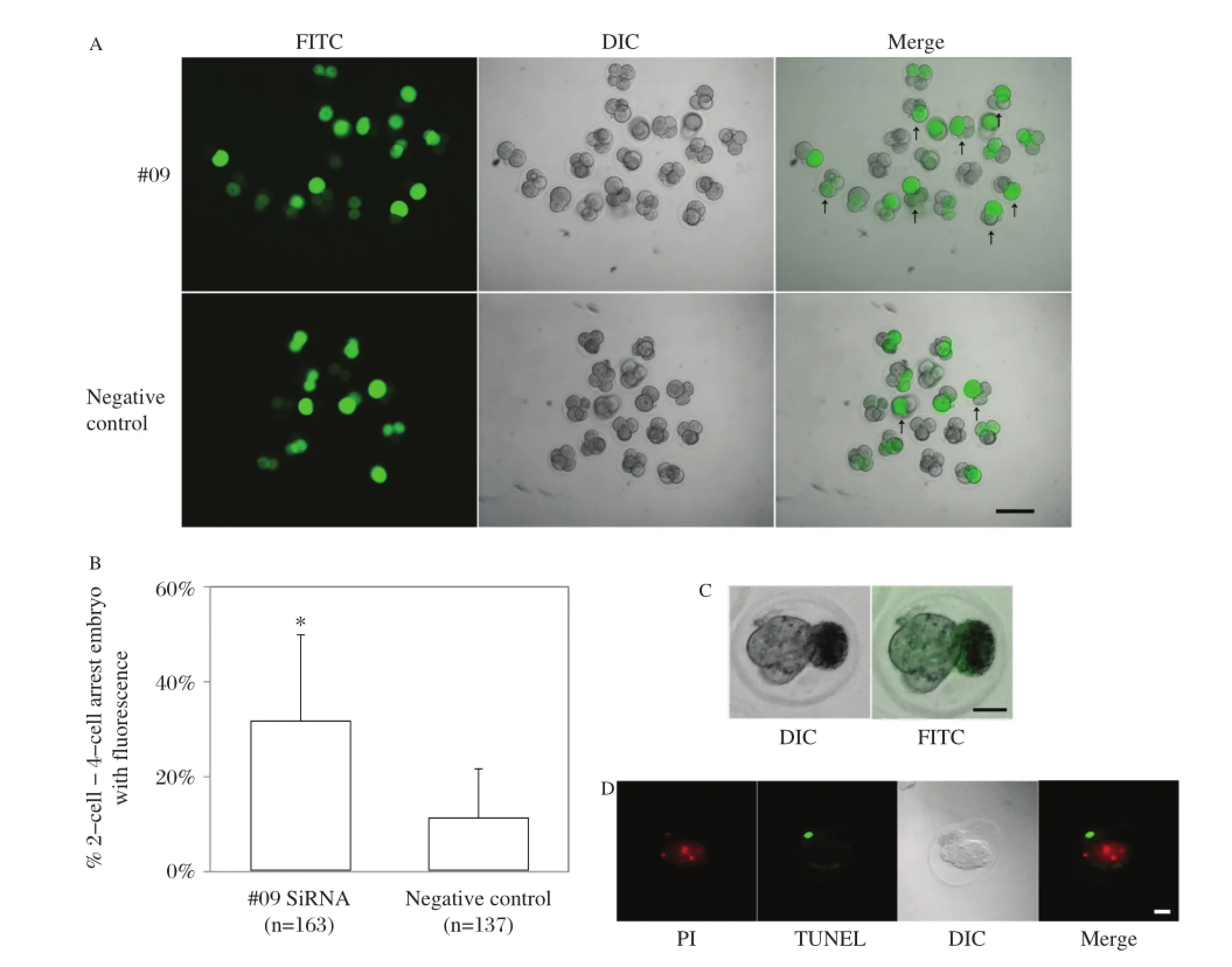
Fig.5Single blastomere microinjection of 2-cell embryo withDdx3xsiRNA induced cell cycle arrest.A:Representative images of embryo development in#09 siRNA and negative control after single blastomere injection with Ddx3x siRNA,indicated by mixed FITC.Green,FITC; white,DIC.The arrow indicats the cell cycle arrest embryo with FITC.Scale bar=100 μm.B:The number of cell cycle arrest embryos was counted and calculated in the#09 siRNA and negative control groups(n=163 and 137 in#09 siRNA and negative control,respectively).C(a):Representative sample for subsequent arrested development of 2–cell to 4–cell embryos,the monoplast with FITC was the blastomere injected by Ddx3x siRNA. C(b):TUNEL staining of the same embryo which showed that the RNAi blastomere was apoptotic when the normal blastomere developed to the morula stage.Scale bar=20 μm.PI,propidium iodide.
When we performed microinjection of Ddx3x siRNA mixed with FITC into the single blastomere of 2–cell embryos during the 2–cell to 4–cell stage transition,we observed that the RNAi group exhibited a greater number of blastomeres with FITC arrest in monoplasts compared with the negative control,and until the subsequent embryo stage quite a few RNAi blastomeres went on to apoptosis.This is primarily due to DDX3X function in cell cycle control,and apoptosis is a secondary effect of cell cycle arrest. The phenotype arises earlier and is stronger than zygote cytoplasm microinjection of Ddx3x siRNA; thus,we hypothesized that DDX3X is critical for 2–cell to 4–cell embryo development;in the late 2–cell stage, almost all maternal DDX3X has degraded and the embryo starts to express its own DDX3X during 2–cell to 4–cell transition.Zygotic–source DDX3X potentially plays a role in the formation of 4–cell embryos. We hypothesized that since the knockdown of Ddx3x in 2–cell is later compared with the zygote,the knockdown of zygotic Ddx3x transcript is more efficient and thorough;the phenotype appeared earlier and was more obvious.
Our data suggest that the accumulation of p53 is potentially the cause of apoptosis in the early embryo; this is in concordance with previous studies.The PI3K–AKT pathway is an important mechanism that main–tains pre–implantation embryo survival(reviewed in[3]).MDM2 is a downstream molecule activated by AKT;the activation of MDM2 is required to keep p53 in a latent state within the embryo[34]and genetic deletion of Mdm2 causes p53–dependent death of early embryos[35].Reduced MDM2 activation leads to failure of p53 degradation,and p53 drives the expression of BAX,which can lead to apoptosis.Knockdown of Wdr74 in zygotes also leads to decreased blastocyst formation and activated p53–dependent apoptosis[36]in which the p53 level is elevated and blocking of p53 function rescues blastocyst formation.
It is well established that in embryos from in vitro fertilization or in vitro culture,there is an elevated level of p53 within the nuclei of blastocysts[37,38],and cultured blastocysts that lacked p53 exhibited a 3–to 4–fold higher rate of successful fetal development following embryo transfer compared with wild type sibling embryos[38];this indicates that the level of p53 is a determinant of normal development of embryos both in the physiological case and in-vitro culture cases,and this level could potentially be considered a marker of an embryo′s quality.Because DDX3X regulates the expression of p53 in early embryos,it can be considered a potential molecular mechanism of unsuccessful early embryo development in vivo and in vitro and may serve as a therapeutic target in assisted reproductive technologies.
In conclusion,the present study demonstrated that knockdown of Ddx3x results in apoptosis or cell cycle arrest in mouse early embryos,and finally leads to the arrest of embryo development;according to the preli–minary results,we suppose the phenotype may be mediated through the p53 pathway;however,the ulti–mate mechanisms mediated by p53 need to be further explored.The data suggest that RNA metabolism–related genes that regulate other genes in transcription and post–transcription are critical during early embryo–genesis.DDX3X plays an essential role in mouse early embryo development,which is a conserved function in accordance with its orthologues in urochordates and several invertebrates.It has been hypothesized that DDX3X in humans may also play a role in early embryogenesis,and it could potentially become a new diagnostic and therapeutic target of impaired blastocyst formation and low birth rate for assisted reproductive technologies.
[1] Schultz RM.Regulation of zygotic gene activation in the mouse.Bioessays 1993;15:531–8.
[2] Nothias JY,Majumder S,Kaneko KJ,DePamphilis ML. Regulation of gene expression at the beginning of mam–malian development.J Biol Chem 1995;270:22077–80.
[3] O′Neill C,Li Y,Jin XL.Survival signaling in the preim–plantation embryo.Theriogenology 2012;77:773–84.
[4] Hara KT,Oda S,Naito K,Nagata M,Schultz RM,Aoki F.Cyclin A2–CDK2 regulates embryonic gene activation in 1–cell mouse embryos.Dev Biol 2005;286:102–13.
[5] Polanski Z,Homer H,Kubiak JZ.Cyclin B in mouse oocytes and embryos:importance for human reproduction and aneuploidy.Results Probl Cell Differ 2012;55:69–91.
[6] Jurisicova A,Latham KE,Casper RF,Varmuza SL. Expression and regulation of genes associated with cell death during murine preimplantation embryo develop–ment.Mol Reprod Dev 1998;51:243–53.
[7] Rosner A,Rinkevich B.The DDX3 subfamily of the DEAD box helicases:divergent roles as unveiled by studying different organisms and in vitro assays.Curr Med Chem 2007;14:2517–25.
[8] Session DR,Lee GS,Wolgemuth DJ.Characterization of D1Pas1,a mouse autosomal homologue of the human AZFa region DBY,as a nuclear protein in spermatogenic cells.Fertil Steril 2001;76:804–11.
[9] Vong QP,Li Y,Lau YF,Dym M,Rennert OM,Chan WY.Structural characterization and expression studies of Dby and its homologs in the mouse.J Androl 2006; 27:653–61.
[10]Rogers GW,Jr.,Komar AA,Merrick WC.eIF4A:the godfather of the DEAD box helicases.Prog Nucleic Acid Res Mol Biol 2002;72:307–31.
[11]Chuang RY,Weaver PL,Liu Z,Chang TH.Requirement of the DEAD–Box protein ded1p for messenger RNA translation.Science 1997;275:1468–71.
[12]Lai MC,Lee YH,Tarn WY.The DEAD–box RNA helicase DDX3 associates with export messenger ribonucleoproteins as well as tip–associated protein and participates in transla–tional control.Mol Biol Cell 2008;19:3847–58.
[13]Soto–Rifo R,Rubilar PS,Limousin T,de Breyne S, Decimo D,Ohlmann T.DEAD–box protein DDX3 associates with eIF4F to promote translation of selected mRNAs.EMBO J 2012;31:3745–56.
[14]Grallert B,Kearsey SE,Lenhard M,Carlson CR,Nurse P, Boye E,et al.A fission yeast general translation factor reveals links between protein synthesis and cell cycle controls.J Cell Sci 2000;113(Pt 8):1447–58.
[15]Liu HY,Nefsky BS,Walworth NC.The Ded1 DEAD box helicase interacts with Chk1 and Cdc2.J Biol Chem 2002; 277:2637–43.
[16]Fukumura J,Noguchi E,Sekiguchi T,Nishimoto T.A temperature–sensitivemutantof themammalian RNA heli–case,DEAD–BOX X isoform,DBX,defective in the tran–sition from G1 to S phase.J Biochem 2003;134:71–82.
[17]Yedavalli VS,Neuveut C,Chi YH,Kleiman L,Jeang KT. Requirement of DDX3 DEAD box RNA helicase for HIV–1 Rev–RRE export function.Cell 2004;119:381–92.
[18]Nekhai S,Jeang KT.Transcriptional and post–transcrip–tional regulation of HIV–1 gene expression:role of cellu–lar factors for Tat and Rev.Future Microbiol 2006;1: 417–26.
[19]Rosner A,Paz G,Rinkevich B.Divergent roles of the DEAD–box protein BS–PL10,the urochordate homologue of human DDX3 and DDX3Y proteins,in colony asto–geny and ontogeny.Dev Dyn 2006;235:1508–21.
[20]Johnstone O,Deuring R,Bock R,Linder P,Fuller MT, Lasko P.Belle is a Drosophila DEAD–box protein required for viability and in the germ line.Dev Biol 2005;277:92–101.
[21]Ma M,Guo X,Wang F,Zhao C,Liu Z,Shi Z,et al. Protein expression profile of the mouse metaphase–II oocyte.J Proteome Res 2008;7:4821–30.
[22]Dai Y,Lee C,Hutchings A,Sun Y,Moor R.Selective requirement for Cdc25C protein synthesis during meiotic progression in porcine oocytes.Biology of Reproduction 2000;62:519–32.
[23]Bi Y,Lv Z,Wang Y,Hai T,Huo R,Zhou Z,et al. WDR82,a key epigenetics–related factor,plays a crucial role in normal early embryonic development in mice. Biol Reprod 2011;84:756–64.
[24]Ma M,Zhou L,Guo X,Lv Z,Yu Y,Ding C,et al. Decreased cofilin1 expression is important for compaction during early mouse embryo development.Biochim Biophys Acta 2009;1793:1804–10.
[25]Cullen BR.Nuclear mRNA export:insights from virol–ogy.Trends Biochem Sci 2003;28:419–24.
[26]Yashiroda Y,Yoshida M.Nucleo–cytoplasmic transport of proteins as a target for therapeutic drugs.Curr Med Chem 2003;10:741–8.
[27]Soto–Rifo R,Ohlmann T.The role of the DEAD–box RNA helicase DDX3 in mRNA metabolism.Wiley Interdiscip Rev RNA 2013.
[28]Stommel JM,Marchenko ND,Jimenez GS,Moll UM, Hope TJ,Wahl GM.A leucine–rich nuclear export signal in the p53 tetramerization domain:regulation of subcellu–lar localization and p53 activity by NES masking.EMBO J 1999;18:1660–72.
[29]Chen L,Shao C,Cobos E,Wang JS,Gao W.4–(methyl–nitrosamino)–1–(3–pyridyl)–1–butanone[corrected] induces CRM1–dependent p53 nuclear accumulation in human bronchial epithelial cells.Toxicol Sci 2010;116: 206–15.
[30]Lai MC,Chang WC,Shieh SY,Tarn WY.DDX3 regu–lates cell growth through translational control of cyclin E1.Mol Cell Biol 2010;30:5444–53.
[31]Berthelot K,Muldoon M,Rajkowitsch L,Hughes J, McCarthy JE.Dynamics and processivity of 40S ribo–some scanning on mRNA in yeast.Mol Microbiol 2004; 51:987–1001.
[32]Lee CS,Dias AP,Jedrychowski M,Patel AH,Hsu JL, Reed R.Human DDX3 functions in translation and inter–acts with the translation initiation factor eIF3.Nucleic Acids Res 2008;36:4708–18.
[33]Geissler R,Golbik RP,Behrens SE.The DEAD–box heli–case DDX3 supports the assembly of functional 80S ribo–somes.Nucleic Acids Res 2012;40:4998–5011.
[34]Jin XL,Chandrakanthan V,Morgan HD,O′Neill C. Preimplantation embryo development in the mouse requires the latency of TRP53 expression,which is induced by a ligand–activated PI3 kinase/AKT/MDM2–mediated signaling pathway.Biol Reprod 2009;80: 286–94.
[35]Jones SN,Roe AE,Donehower LA,Bradley A.Rescue of embryonic lethality in Mdm2–deficient mice by absence of p53.Nature 1995;378:206–8.
[36]Maserati M,Walentuk M,Dai X,Holston O,Adams D, Mager J.Wdr74 is required for blastocyst formation in the mouse.PLoS One 2011;6:e22516.
[37]Chandrakanthan V,Chami O,Stojanov T,O′Neill C. Variable expressivity of the tumour suppressor protein TRP53 in cryopreserved human blastocysts.Reprod Biol Endocrinol 2007;5:39.
[38]Li A,Chandrakanthan V,Chami O,O′Neill C.Culture of zygotes increases TRP53[corrected]expression in B6 mouse embryos,which reduces embryo viability.Biol Reprod 2007;76:362–7.
Received 10 April 2013,Revised 06 June 2013,Accept 04 August 2013,Epub 03 March 2014
?Corresponding author:Dr.Jiahao Sha,State Key Laboratory of Reproductive Medicine,Nanjing Medical University,Hanzhong Road 140,Nanjing,Jiangsu 210029,China.Tel:+86–025–86862908,E–mail:shajh@njmu.edu.cn.
The authors reported no conflict of interests.
?2014 by the Journal of Biomedical Research.All rights reserved.
10.7555/JBR.27.20130047
 THE JOURNAL OF BIOMEDICAL RESEARCH2014年4期
THE JOURNAL OF BIOMEDICAL RESEARCH2014年4期
- THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Evaluation of quantitative variation of secondary metabolites in Bergenia ciliata (Haw.)using high performance thin layer chromatography
- Comparing a non-invasive hemodynamic monitor with minimally invasive monitoring during major open abdominal surgery
- Morphological MRI and T2 mapping of cartilage repair tissue after mosaicplasty with tissue-engineered cartilage in a pig model
- Schistosoma japonicum infection induces macrophage polarization
- Luteolin prevents uric acid-induced pancreatic β-cell dysfunction
- IRE1αknockdown rescues tunicamycin-induced developmental defects and apoptosis in Xenopus laevis