
Proximity association in polygynous western black crested gibbons(Nomascus concolor jingdongensis): network structure and seasonality
2013-12-17 09:17:04Zhen-HuaGUAN,BeiHUANG,Wen-HeNING等
Zoological Research 2013年1期
Spatial distance between individuals exists in all group living animals; however, variances occur among different species and among individuals within the same group. It is reported that spatial distance among individuals is correlated with social relationships (Sade,1965, 1972) and social organization (Kummer, 1968);therefore spatial proximity between group members is important for studying social structure. Early research reported on a correlation between inter-individual distance and social relationship in Macaca mulatta (Sade,1965), with further studies using spatial data to represent social structure in different primates (e.g. Symphalangus syndactylus: Chivers, 1971; Papio cynocephalus and Saimiri sciureus: Fairbanks, 1976). In recent years,spatial distance has also been used in species with different social structures, such as multi-male-multifemale groups (e.g. Macaca mulatta: Sherman, 1980;Macaca nemestrina and Macaca arctoides: Weigel, 1980;Macaca fuscata fuscata: Corradino, 1990) and multilevel society groups (e.g. Theropithecus gelada: Snyder-Mackler et al, 2012; Papio hamadryas: Schreier &Swedell, 2012; Swedell & Plummer, 2012;Rhinopithecus roxellana: Zhang et al, 2012).1
Social structure in animal groups can be affected by many factors, such as kinship (Furuichi, 1984;Matsumura & Okamoto, 1997; Silk, 2002), age classes(de Waal & Luttrell, 1986), resource holding potential(Barrett & Henzi, 2006), and resource availability (Henzi et al, 2009). A number of studies have also investigated the relationship between ecological factors and social behavior (Emlen & Oring, 1977; Terborgh, 1986; Sterck et al, 1997; Kappeler & Van Schaik, 2002) and shown that social relationships may change in different seasons(Shimooka, 2003) and habitats (Pruetz & Isbell, 2000).However, compared to numerous studies on the effects of seasonal ecological factors on animal behavior (Dunbar,2002; Dunbar et al, 2009), such as activity budgets (e.g.Theropithecus gelada: Dunbar, 1992; Ateles spp.:Korstjens et al, 2006; Nomascus concolor: Fan et al.,2008; Eulemur fulvus fulvus: Sato, 2012; Nomascus nasutus: Fan et al, 2012) and range behavior (e.g.Theropithecus gelada: Dunbar, 1998; Symphalangus syndactylus: Kim et al, 2011), how social relationships are affected by ecological factors is not well understood,with very few studies conducted on small apes.
Social relations are not easily investigated(Whitehead, 1997), and the development of appropriate measures for spatial proximity can express qualitative variation and assess the social structures (Cairns &Schwager, 1987; Whitehead, 1995; Ramos-Fernández et al, 2009). Early research on spatial distance in siamangs(S. syndactylus: Chivers, 1971) considered spatial relationship as an important measure for group cohesion.However, few studies have addressed social relationships or spatial proximity in gibbons. While most gibbon species live in small monogamous groups (Leighton,1987), recently several gibbon species have been found to live with flexible group composition (Brockelman,2009). Lappan (2007) used spatial distance to study social relationship among adults in multi-male siamang groups (S. syndactylus). Stable multi-female groups only exist in the black crested gibbon (Nomascus concolor:Jiang et al, 1999; Fan et al, 2006), Hainan gibbon (N.hainanus: Zhou et al, 2005) and Cao Vit gibbon (N.nasutus: Fan et al., 2011). Polygynous groups generally have more individuals (Jiang et al, 1994b; Fan et al, 2006;Zhou et al, 2008), and studying social relationships in such groups (e.g. N. concolor: Fan & Jiang, 2009) can aid understanding of the social system evolution mechanism in gibbons. However, no research on the influence of ecological factors to spatial proximity in gibbons has been conducted to date, though Fan et al(2008, 2012) found black crested gibbons and Cao Vit gibbons were more likely to stay together in cold seasons during the night.
Our aim was to study proximity associations in a polygynous group of western black crested gibbon in Mt.Wuliang, central Yunnan, and assess the impacts of ambient temperature and rainfall on seasonal variance.We also considered the influence of anthropogenic disturbance on proximity among group members.December 2011. Our study group was habituated in December 2010 after thirty months of previous following.The group was composed of seven individuals: one adult male, two adult females, one sub-adult male, two juveniles, and one infant (Table 1). All individuals were recognized by the observers according to their facial features and body size. We named the individuals of the group following Guan et al (submitted) (Table 1). InR was born in November 2009 and was still dependent on its mother. JuI was born in 2008 and JuR was born in 2006, both were fully independent.

Table 1 Composition of the study group
METHODS
Study site and subjects
We carried out the study at Dazhaizi (N24°21’,E100°42’) on the western slopes of Mt. Wuliang, in Jingdong County, central Yunnan, China. We used a HOBO pro RH/Temperature Data Logger (RG3-M) to record the temperature every two minutes and rainfall continuously in the study area from January 2011 to
Data collection
We spent approximately 20 days per month in the field. Every morning the group was located by listening for their morning duets, going directly to their sleeping sites, or by visiting their food patches. Once the group was located, we followed it as long as possible and used scan sampling at 5-min intervals to collect data on the spatial distance among all independent individuals, with spatial proximity defined as individuals within 1 m of each other at the time of scan sampling. For collection of distance data, we estimated distance between any two trees by eye and confirmed it using a direct-reading optical range finder (OLC 600XV) in 2010. We achieved a distance estimation of <10 m with error <1 m and 10-20 m estimation with a ca. 2 m error. We then collected systematic data between January 2011 and December 2011.
We considered one observing day as effective only if the group was followed for more than 5 h in that single day, and only data collected on effective observing days were included in the following analysis.
Data analysis
Simple ratio association index (hereafter SRI) was used to study pairwise associations. The SRI was calculated using the number of scans in which two individuals were seen in proximity divided by the total number of scans during which the dyad was in proximity and was separated (Cairns & Schwager, 1987):
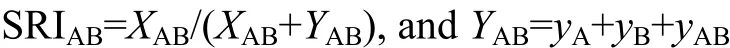
In the formula, XABstands for the number of scans in which A and B were seen in proximity while YABmeans they were not; yAand yBmean the number of samplings when only A was located or only B was located separately; and yABrepresents the number of scans in which both A and B were located but were not in proximity. For each pair, total observation time varied among months. To avoid the influence of different sampling size, we first calculated monthly SRI and then averaged them to obtain annual mean SRI for every dyad.
Social network analysis (hereafter SNA) is a useful tool for studying social structure in different primates(Kasper & Voelkl, 2009; Whitehead, 2009). In this paper,SNA was used for studying the proximity network,which was built in Ucinet 6.0 (Borgatti et al, 2002). We first constructed a weighted proximity network using annual SRI of each dyad, which represented the presence or absence as well as the frequency of proximity distance.We used group density as a measure of group cohesion,which was defined as the number of observed dyads with proximity divided by the number of possible dyads according to the number of individuals in the network(Hanneman & Riddle, 2005). To highlight the more important dyads in the network, we then defined preferred proximity dyads as those with SRI higher than or equal to the mean SRI of all possible dyads(Kanngiesser et al, 2011). We built an unweighted network by the transformation of SRI into binary values with 1 standing for the preferred proximity dyads and 0 standing for the non-preferred ones, i.e., only preferred proximity dyads were retained in this network. This approach has not been well validated in former studies,so we also used quadratic assignment procedure (QAP)to test whether the unweighted network adequately reflected the structure of the weighted network.Bootstrap replicates were used to compare the density of preferred proximity networks to a theoretical fully connected network (density=1) to investigate whether preferred proximity was distributed randomly.
We conducted hierarchical cluster analysis in Socprog 2.4 (Whitehead, 2009) to assess the associations among hierarchically formed clusters. Cophenetic correlation coefficient indicated the fitness of the dendrogram as representative of the observed data.Cophenetic correlation coefficients>0.8 (Whitehead,2009) indicate a good match. We used a paired sample test to investigate whether the adult male had different proximity association with the two females and their offspring by comparing their monthly SRI. Kolmogorov-Smirnov test was used for normality before we made comparisons of different dyads. Paired sample test was also used to compare the proximity association of one female and her two offspring of different ages. To access the trend of proximity association change of motheroffspring dyads, we used multiple linear stepwise regression to test the impact of month (i.e. when the juveniles or infant matured) and environment factors(temperature and rainfall). To describe the variance of seasonal social proximity, we calculated the mean SRI of the group in twelve months, and used linear stepwise regression to test the impact of temperature and rainfall.Because the group was newly habituated in January 2011 and was not as well observed as in the other eleven months, we removed the January data and estimated the influence of weather again. All statistical analyses were conducted via SPSS 16.
RESULTS
The mean temperature during the study period was 15.7 °C, with the lowest mean in January (10.1 °C) and the highest in June (19.2 °C). Precipitation was 1 793 mm from January 2011 to December 2011, with an obvious wet season from May to October during which 84% of the rainfall occurred (Table 2).
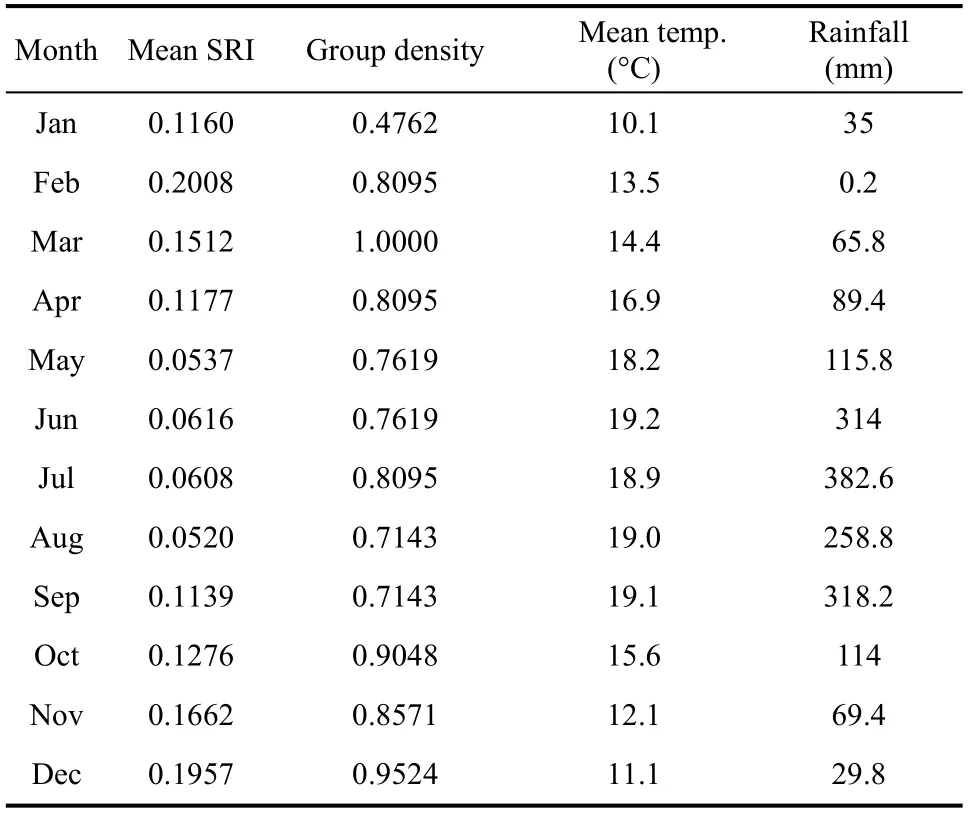
Table 2 Seasonal variation of monthly mean SRI and group density from January to December in 2011
During our study period, 5 229 scans were made and 31 013 distance data were collected. Group density varied monthly. The group was cohesive (group density>0.7) in all months except January (lowest density value of 0.48,Table 2). However, annual group density was 1 (fully connected) for the weighted network as all possible dyads were observed in proximity (n=21). The preferred proximity (unweighted) network well reflected the structure of the weighted network (QAP, R=0.609,P=0.001). A preferred proximity relationship only existed in about half of all possible dyads (density was 0.4762 for this network, i.e. 10 preferred proximity dyads in 21 possible pairs) and preferred proximity was not distributed evenly (Bootstrap test: z = ?3.0755,P=0.0002). The individual SAM2 exhibited no preferred proximity with other members, which confirmed his peripheral position (Figure 1).
Using hierarchical cluster analysis, we identified four subgroups with a cophenetic correlation coefficient of 0.98: the two females with their own offspring, the adult male (AM2), and the sub-adult male (SAM2).Results showed that AM2 preferred proximity with AFR and her offspring more often than with AFI and her offspring (Paired sample test: AM2-AFR vs AM2-AFI,df=11, t=3.048, P=0.011; AM2-InR vs AM2-JuI: df=11,t=3.230, P=0.008: AM2-JuR vs AM2-JuI, df=11, t=2.274,P=0.044) (Figure 2).
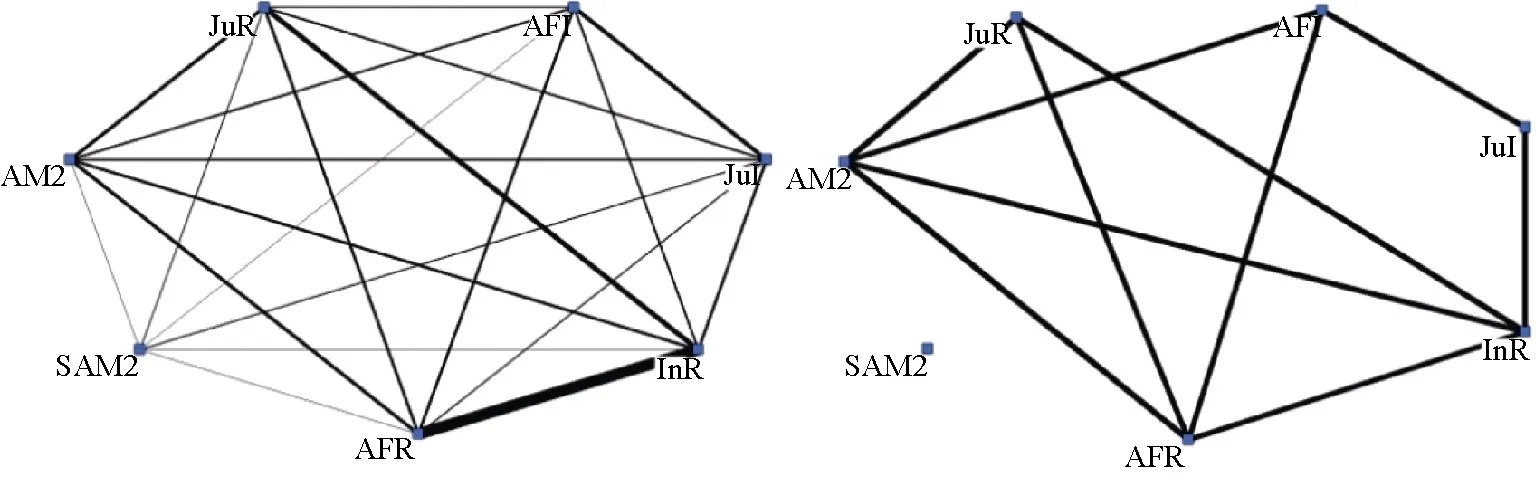
Figure 1 Weighted (left) and unweighted (right) social network of proximity (using Ucinet 6.0)Nodes represent individuals and lines represent the presence of association between individuals. In the weighted network, the thickness of the line represents the frequency of proximity position.
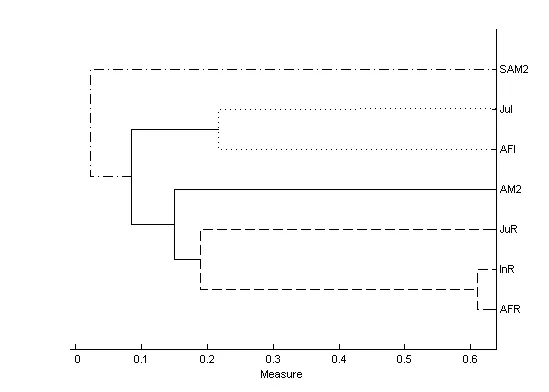
Figure 2 Dendrogram of proximity associations between individuals created in SCOPROG 2.4 (Whitehead,2009)Labels on the right axis represent the name of individuals. Four subgroups are identified by different line types.
Three mother-offspring pairs were compared, and all SRI data were normalized distributed (Kolmogorov-Smirnov test: AFR-InR, n=12, P=0.657; AFR-JuR, n=12,P=0.352; AFI-JuI, n=12, P=0.943). The SRI between mother and dependent offspring (AFR-InR) was significantly higher than between mother and independent offspring (AFR-JuR) (Paired samples test:df=11, t=7.161, P=0.000), but no significant variance was observed between two independent offspring (Paired samples test: AFR-JuR vs AFI-JuI, df=11, t=?2.048,P=0.065). Stepwise regression showed that the proximity of the dependent infant with their mother was only affected by age, that is, proximity decreased with age(InR-AFR, F1,10=36.992, r2=0.787, P=0.000, Figure 3).However, the SRI of fully independent offspring and their mother was significantly affected by mean temperature (JuI-AFI, F1,10=6.205, r2=0.383, P=0.032)and rainfall (JuR-AFR, F1,10=5.014, r2=0.334, P=0.049).The group’s mean SRI changed monthly (Table 2) and ambient temperature significantly influenced proximity association (F1,10=14.404, r2=0.591, P=0.004). After removing January data, the impact of temperature increased (F1,9=42.315, r2=0.825, P=0.000). The SRI increased in the cold months, with the highest in February and December (0.20). The group maintained low proximity association while the mean temperature was high from May to August, with the lowest SRI (0.05)appearing in May and August (Figure 4).
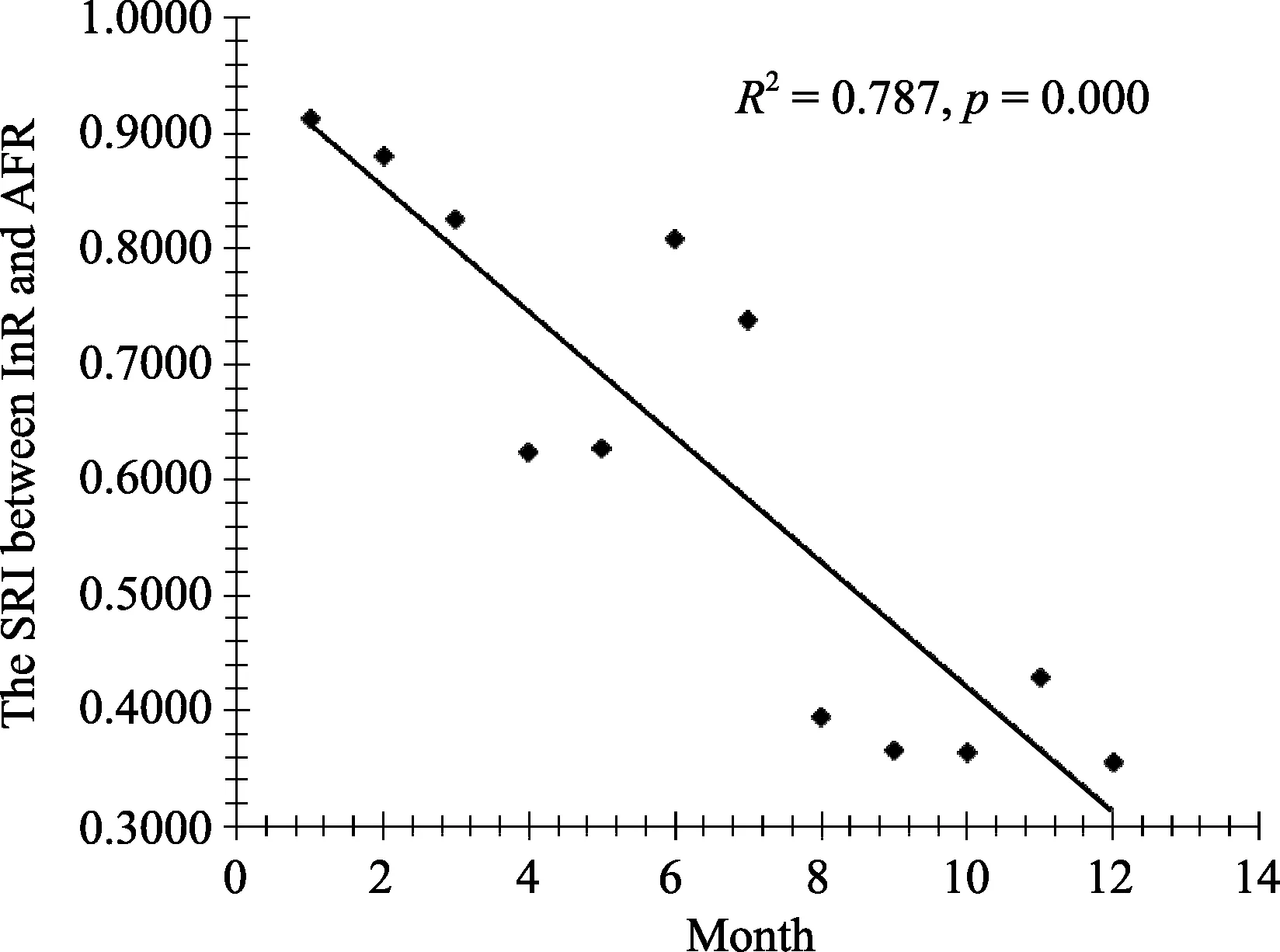
Figure 3 SRI between mother and dependent offspring(AFR and InR)
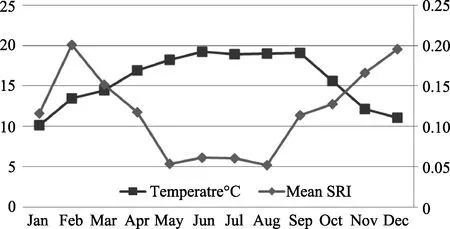
Figure 4 Correlation between mean temperature and group mean SRI
DISCUSSION
Social structure of a polygynous group of gibbons
Kasper & Voelkl (2009) investigated social cohesion in 70 primate species, and found a mean density of 0.75 (n=70, range 0.49?0.93). Gibbons live in small groups. Although the claim of nuclear family groups may not be correct in many gibbon species, and adult replacement as well as extra-group copulation have been observed in our study group (Fan et al, 2010;Huang et al, accepted), this group still showed a fully connected network (density=1), which meant all individuals directly connected with each other and represented very high social cohesion. This may be how gibbons maintain group stability, especially when paternity is uncertain. However, the proximity association between different individuals was uneven.Only ten dyads exhibited preferred proximity, and the sub-adult (SAM2) obviously exhibited peripheral clustering in the study group, as has been shown in previous research (Fan & Jiang, 2009). Four clustering units were identified and kinship was a good explanation for the observed pattern (Furuichi, 1984; Matsumura &Okamoto, 1997), as females preferred to keep proximity with offspring. Even among young gibbons, individuals also maintained proximity with individuals of the same matrilineal line. When the dependent offspring grew older, the proximity between mother and offspring declined, as observed in saki monkeys (Pithecia pithecia:Thompson & Norconk, 2011).
In our study group, the most notable characteristic was that the adult male had different proximity association with the two adult females. This differs from previous research, which reported that two females maintained similar spatial distance from the adult male in the neighboring group (Fan & Jiang, 2009). This variance may be due to different analysis methods as we defined 1 m as a proximity distance while Fan & Jiang(2009) used mean spatial distance for measuring the relationship. Considering the group history and different reproductive status of the two females (infant-carrying or not), we can give two possible explanations for our results.
Previous research has shown infanticide affects social structure (Sterck et al, 1997). Reichard (2003)argued that infanticide is a potential threat for newborns and also concluded that infanticide avoidance could drive the evolution of monogamy in gibbons, suggesting females also face infanticide stress. In 2008, AFI and her newborn infant (JuI) immigrated into the study group(Huang et al, accepted; Guan et al, submitted). This female was never seen previously in the study area, and the paternity of the infant was unknown. Such circumstances increase the risk of infanticide. Although infanticide has never been reported in gibbons, with the possible exception of one case in Khao Yai (Reichard,2003), we considered that infanticide avoidance may be one reason for AFI to limit proximity with AM2. The adult male AM2 grew up in the neighboring group and immigrated into our study group in 2007, replacing the original adult male (Fan et al, 2010). Thus we cannot be sure whether AM2 is or is not the biological father of any young within the group (InR and JuI). Genetic data would help solve this uncertainty.
Alternatively, infants may affect the relationship between males and females because male proximity is important to protect a dependent infant from predation(Dunbar & Dunbar, 1980). This has been reported in small primates such as titis monkeys (Callicebus discolor:de Luna et al, 2010) and saki monkeys (Thompson &Norconk, 2011), as well as in white-handed gibbons(Hylobates lar: Reichard, 2003) and siamangs(Symphalangus syndactylus: Lappan, 2008). In our study area, gibbon infants face predation from eagles (Spilornis cheela) (personal observation), and obviously dependent offspring (InR) would most likely require protection.Although prevention from predation is considered a more efficient strategy in monogamous than in polygynous groups (Sommer & Reichard, 2000), we found that the adult male kept proximity with the female carrying dependent offspring (AFR) in the polygynous group.
Long term observation is necessary to determine which explanation is more reasonable. If the male maintains greater proximity with the infant-carrying female, the latter protection hypothesis can be proved; if the proximity of AFI and AM2 increases as JuI matures,the infanticide-avoidance hypothesis would be preferred;if the male maintains proximity to AFR, the impact of individual preference or/and sibship among individuals may be considered; or, it is also possible that the male maintains proximity to one female over another randomly among years.
Seasonal variance of proximity
Temperature influences on group proximity
Many studies have reported on the influences of ambient temperature on animal behavior (e.g.Theropithecus gelada: Dunbar, 1998; Symphalangus syndactylus: Kim et al, 2011; Eulemur fulvus fulvus: Sato,2012; Nomascus concolor jingdongensis: Fan et al,2008). In our study, we found that temperature significantly influenced group proximity, with individuals spending more time in spatial proximity when the temperature was low. This phenomenon was likely a response to thermoregulatory needs, which is an important ecological force (Hill et al, 2004). Gibbons in Mt. Wuliang inhabit the northern extreme of Hylobatidae habitat and are at high altitude with extreme seasonality of temperature and rainfall, resulting in great challenges associated with thermoregulatory costs. Previous studies have shown that both black crested gibbons in Mt.Wuliang (Fan & Jiang, 2008) and Cao Vit gibbons in Guangxi huddled together in their sleeping places in cold months while they slept separately or apart in the warmer months (Fan et al, 2012). Our study showed that gibbons also increased time in proximity through the day time in cold months.
According to our results, group proximity decreased regularly from the dry season to the rainy season and reached the lowest point from May to August. This may relate to the seasonal time budget in the study subjects.Food availability increased in the rainy season and the gibbon group increased feeding time and decreased resting time (Huang, 2011). The gibbons seldom stayed at proximity distance when feeding, singing, and moving,with most proximity/touching occurring during resting time. Therefore, when the group decreased resting time,they also decreased the time spent in proximity distance.
Human activity influences on group proximity
Anthropogenic disturbance might also influence group proximity. In social animals, an individual’s fitness depends partly on the quality of relationships with others(Schülke et al, 2010; Silk et al, 2010; Seyfarth & Cheney,2012). Good social relationships provide partners with direct benefits such as grooming and support in aggressive conflicts, or increase tolerance near resources and, thus, decrease competition (Silk, 2007; Massen et al,2010; Seyfarth & Cheney, 2012). However, these benefits might disappear when the group meets severe disturbance.
In a previous study, Fan & Jiang (2010) found gibbon behavior changed in high human disturbed areas.Most human activity (livestock grazing) in the study area was found at low altitude (Fan & Jiang, 2010). The low proximity from May to August may be due to high frequency of human disturbance. In those months, more food resources were available at low altitude and mushroom collecting occurred more frequently, thus gibbons had to increase moving time and keep alert.Gibbons may increase distance between individuals, or even move in different directions when avoiding human disturbance (Jiang et al, 1994a). Although gibbons spent more time in the low area from November to March,human disturbance was lower in those months. After August, gibbons spent most of the time in the high altitude area (over 2 300 m), where human activity was rarely found.
Acknowledgements: We carried out this study at the Black Crested Gibbon Monitoring Station at Mt.Wuliang. We acknowledge the staff from the Jingdong Nature Reserve Management Bureau for their valuable support. We acknowledge our field assistants, Mr. Shi-Ming XIONG and Mr. Ye-Kun LIU, for their kind help.

- Zoological Research的其它文章
- Blockage of glucocorticoid receptors during memory acquisition,retrieval and reconsolidation prevents the expression of morphineinduced conditioned place preferences in mice
- Seasonal variations in the energy budget of Elliot’s pheasant(Syrmaticus ellioti) in cage
- Seed caching and cache pilferage by three rodent species in a temperate forest in the Xiaoxinganling Mountains
- 中國鳥類亞種新記錄——黑冠黃鵯
- 虹鱒LECT2的酵母表達、純化及生物活性分析
- 蛇毒抗菌肽OH-CATH對大腸桿菌引起家兔泌尿系感染的保護作用