
Quantification of Oligodendrocyte Progenitor Cells in Human and Cat Optic Nerve:Implications for Endogenous Repair in Multiple Sclerosis*
2010-06-06 10:57:24ALISONJENNINGSANDWILLIAMCARROLL
神經(jīng)損傷與功能重建 2010年5期
ALISON JENNINGSAND WILLIAM CARROLL
1.School of Pathology and Laboratory Medicine,University of Western Australia,Nedlands,Western Australia,Australia;2.Department of Neurology,Sir Charles Gairdner Hospital,Perth,Western Australia,Australia
INTRODUCTION
Animal models have demonstrated complete repair of demyelination,effected by replacement oligodendrocytes arising from the division and immigration of oligodendrocyte progenitor cells(OP)(Blakemore and Keirstead,1999;Carroll and Jennings,1994;Carroll et al.,1998).In contrast,a feature of multiple sclerosis(MS)is the eventual predominance of chronically demyelinated lesions,making enhancement of endogenous repair an appealing strategy for improving disease outcome(Baer et al.,2009;Calza et al.,2005;Gallo and Armstrong,2008;Lubetzki,2008;Omari et al.,2009;Rist and Franklin,2008).An essential pre-requisite is determining the baseline capacity of the human CNS to generate remyelinating oligodendrocytes.Pivotal to this is the reliable identification of OP in human tissue and their quantification relative to oligodendrocytes(Staugaitis and Trapp,2009).
Although putative OP have been identified using the surface antigenic markers,NG2 and PDGFα R,demonstration is restricted to suitably preserved,frozen tissue(Reynolds et al.,2002;Staugaitis and T rapp,2009).Doubt has also been cast on whether all NG2+neuroglial cells(NG2cells)are OP.With NG2cells found in comparable numbers in nonmyelinated and myelinated regions(Chittajallu et al.,2004;Dawson et al.,2003),involved in glial scar formation(Jones et al.,2002;Mc-Tigue et al.,2006;Tan et al.,2005)and shown to have associations with synapses and nodes of Ranvier(Belachew and Gallo,2004;Bergles et al.,2000;Butt et al.,1999;Hamilton et al.,2009),opinion has been divided between those who consider that all NG2cells are“polydendrocytes”with the potential to act as OP(Dawson et al.,2003;Nishiyama et al.,2009;Polito and Reynolds,2005)and those who believe that OP comprise only a fraction of NG2cells,the majority of which belong to a separate class of differentiated neuroglia termed “complex cells”or “synantocytes”(Butt et al.,2002,2005;Greenwood and Butt,2003;Horner et al.,2002;Ligon et al.,2006;Peters,2004;Wigley and Butt,2009).Clearly,the latter scenario could have major implications for strategies to maximise endogenous repair in MS that are premised on robust proliferative responses by “numerous” or“abundant” (NG2+)OP(T anaka et al.,2003;Wilson et al.,2006;Yang et al.,2006).Therefore,there is a need to determine whatneuroglial cell type/s express NG2,including the possibility of subpopulations with functional differences,particularly the capacity to act as OP to effect remyelination.The essential endpoint is to know how many OP are available for oligodendrocyte replacement in the normal adult human CNS.
This study has addressed these issues in the optic nerve(ON),which is commonly affected in MS,provides a structurally simplified model of CNS white matter(WM),and has been the source for much of our knowledge of the oligodendrocyte lineage in vitro and in vivo(Bolton and Butt,2005;Ffrench-Constant and Raff,1986;Fulton et al.,1992;Hogan and Zimmerman,1968).Historically,the cat has been chosen for studies of visual system electrophysiology as well as response to demyelination(Bunge et al.,1960;Carroll et al.,1985;Cook,1979;Rose et al.,1997).In the experimental cat optic nerve(ECON)model of demyelination and remyelination,microinjection of antisera to galactocerebroside causes the demise of oligodendrocytes and OP within a focal lesion(Carroll et al.,1985).Immunohistochemistry(IHC)correlated with ultrastructure has revealed that successful lesion repair by remyelinating oligodendrocytes is the result of a stepwise maturational progression from the progeny of peri-lesion activated OP(aOP)(Carroll and Jennings,1994;Carroll et al.,1990,1998).The subsequent description of OP in normal adult cat ON as a single neuroglial population of relatively low abundance,together with evidence from autoradiographic tracking that aOP division was symmetric,raised the possibility that replenishment of OP might be compromised in protracted or large-scale demyelination(Carroll et al.,1998;Jennings et al.,2002).
Although human tissue studies have more relevance to MS(Friese et al.,2006;Miron et al.,2007),relatively poor tissue preservation associated with postmortem autolysis has challenged accurate cell identification(Peters,2002).In this study,however,results obtained in perfusionfixed normal cat ON were able to be directly validated in short postmortem interval(pmi)human ON and subsequently in formalin-fixed paraffinembedded(FFPE)subsamples.Demonstration of the OP surface antigen markers NG2 and PDGFα R,in correlation with the oligodendrocyte lineage nuclear transcription factors,Olig1 and Olig2 and previously characterized intracellular markers(Jennings and Carroll,1999;Jennings et al.,2002)has resulted in an antigenic phenotype for NG2cells in frozen,paraffin,and resin sections.NG2cells were found to comprise a single neuroglial class without detectable subpopulations.Although morphologically complex,all NG2cells appeared to have OP capability and could appropriately be called polydendrocytes(Nishiyama,2007;Nishiyama etal.,2002,2009).Importantly,NG2cells were not abundant,warranting consideration in neuroprotection and enhancement of endogenous repair in MS.
MATERIALS AND METHODS Tissue Samples and Section Preparation Feline
Samples of normal,4%paraformaldehyde-perfused,cat ON,and spinal cord(SC),provided by Dr Suomo Ghosh(Ring et al.,2004),were postfixed for 6 h and then processed for paraffin or OCT embedding(cryoprotection in 30%sucrose;snap freezing in liquid nitrogen-cooled isopentane).Araldite-embedded ON comprised normal controls and blocks containing an early(2 day postinjection)demyelinative lesion generated during previous studies(Carroll and Jennings,1994;Carroll et al.,1990,1998).
Human(Table 1)
Fresh low-pmi(<12 h)intraorbital ON samples from corneal transplant donations to the Lions Eye Bank,Perth,Western Australia(Sir Charles Gairdner Hospital HREC approval no.2004~055)were immersed in a zinc salt-based fixative(hereafter,called zincfix)(Beckstead,1994;Lykidis et al.,2007).Transverse and longitudinal orientation blocks were then either embedded in OCT for frozen sectioning or in paraffin wax following postfixation in either 10%buffered formalin(for up to 2 weeks)or zincfix.
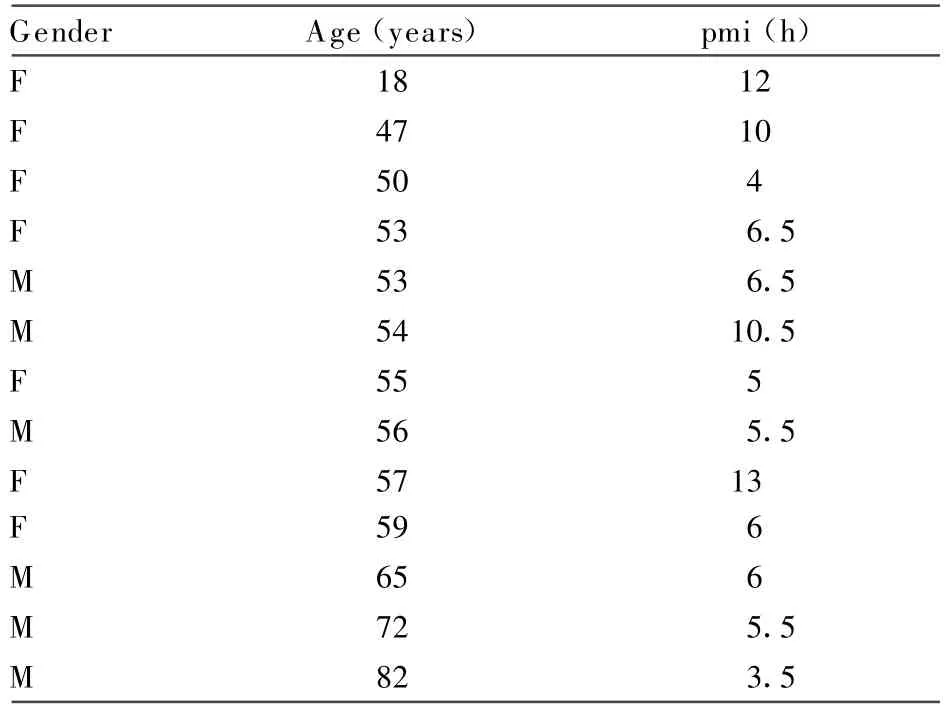
TABLE 1.Detailsof Fresh Human Optic Nerve Samples
Multiblocks
Multiple pieces of frozen or paraffin-processed cat and/or human tissue embedded togetherto form multiblocks maximized information from individual sections and provided internal immunostaining standardization.For resin-embedded ON,multiple sections were collected on individual slides.
Section preparation
Blocks were serially sectioned onto silanated slides,at 1 μ m for resin,2 ~ 3 μ m for paraffin,and 3 ~ 5 μ m or 10 μ m for frozen blocks.Before immunostaining,resin sections were deplasticized and etched as described previously(Jennings et al.,2002);frozen sections were air dried at room temperature(RT)for 1 h,and paraffin sections were dewaxed and hydrated.
IHC(Table 2)
For antigen retrieval(AR),slides immersed in 0.05%citraconic anhydride(pH 7.4)(Namimatsu et al.,2005)were microwaved for 30 min at 100℃(BioRad H2500 Microwave Processor)and then cooled to RT.Nonspecific protein binding wasblocked with 10%normalhorse serum(NHS).Sections were incubated with primary antibodies(Table 2)diluted in phosphate-buffered saline at 37℃,RT or 4℃,for durations tailored to the individual antibody/ies.Endogenous peroxidase activity was blocked with 0.3%hydrogen peroxide for 15 min.After single primary antibody reactions or where two antibodies raised in the same species but to separate antigenic entities were mixed and reacted together(e.g.,rabbit antibody to astrocyte cytoplasm and oligodendrocyte nuclei,respectively),EnVision(Dako)was applied for 40 min at RT,and staining revealed with DAB(diaminobenzidine,liquid DAB1;Dako).In multiple stain protocols,primary antibodies applied as a cocktail were separately revealed via species-or immunoglobulin isotype-specific secondary link reagents,followed by appropriate chromogen and counterstain combinations to maximize color contrast(T able 2).For instance,to achieve dual staining,one primary antibody was labeled with horse radish peroxidise and revealed with DAB(brown);the other with alkaline phosphatase,revealed by Fuchsin(red),followed by counterstaining with hematoxylin(blue-purple).Where a third chromogen was required,SG(gray-blue)was used and counterstaining either omitted or kept very light.Staining controls included positive and negative control tissues within multiblocks and primary antibody omission.
Demonstration of NG2 and PDGFα R
To achieve surface antigen staining while protecting tissue integrity,minor variations were made to the basic IHC protocol.AR was not performed.Triton-X was included in the NHS-blocking solution,at 0.3%for frozen sections and at 0.15%for paraffin sections.Before frozen section primary antibody incubation,light nuclear staining with hematoxylin improved nuclearpreservation whileconfirming whether immunopositivity was cellular.For NG2 demonstration,primary antibody incubation was for 5 days at 4℃;secondary antibody incubation was for 90 min.

TABLE 2.Primary Antibodies,Secondary Antibodies,and Chromogens
Imaging and Quantifigcation Bright figeld light microscopy(LM)
LM images up to×100 objective magnifigcation(obj)oil immersion were captured from a Zeiss U-niversal Research Microscope figtted with a Leica DC100 digital camera linked to Leica IM50 software.
Virtual microscopy(VM)
An Aperio T2 ScanScope automated slide scanner(Aperio Technologies,Vista,CA)was used to digitally scan slide-mounted sections in their entirety at×40obj.The resultant ScanScope virtual slide(“.svs”)image could then be viewed,imaged,and manipulated on a computer at magnifigcations from slide overview up to×40.
Quantifigcation
Cells were counted on VM slides within standard area frames that each enclosed 0.2 mm2(standard figeld).Image annotation software recorded all raw cell counts as well as permitting a visual record of the location of counting frames across the face of cross-sections.For each cell type,counts from multiple figelds were averaged for expression as cells per mm2(±standard deviation;SD)and also pooled for expression as relative percentages of the oligodendrocyte lineage,macroglia,or neuroglia(Tables 4~6).In cat ON,the reliability of pooled standard figeld counting wascorroborated by counting full-face cross-section totals of NG2cells,oligodendrocytes,and astrocytes(Table 4).T o control for possible artefactual count differences due to differential tissue density,raw count data comparisons were only made between identically figxated and prepared specimens.To ensure the correct staining designation ofindividual cells within digital slide counting frames,sections were viewed concurrently under the microscope at×100obj.
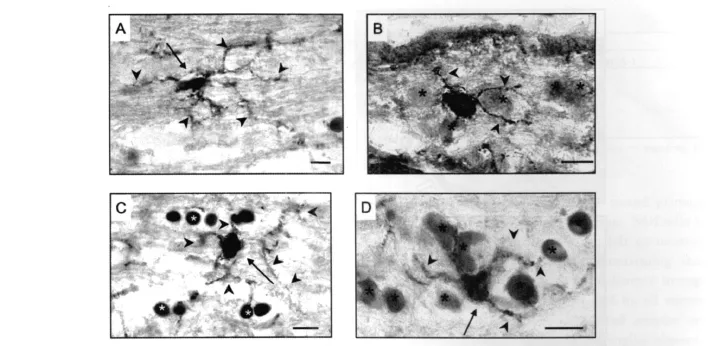
Fig.1. NG2 expression in frozen sections.Cat(A,C-D)and human ON(B);neuroglial cells with surface positivity for NG2 outlining their cell body and processes(NG2cells)were located both alone among the fiber bundles(A,C)and among other neuroglia(asterisks)in intrafascicular and perifascicular rows(B,D).Morphological heterogeneity was evident in terms of nuclear shape and process orientation.A:The majority of NG2cells featured small cell bodies closely surrounding elongated,irregular outline nuclei(arrow),with plentiful,long,mainly polar,fiber-associated processes(arrowheads).B-D:Other NG2cells,both spanning across the fibers(C)and among other neuroglia(asterisks)(B,D)had relatively rounded nuclei(arrows)and more radial processes(arrowheads).Bars=10μ m.
Quantifigcation and Morphological Description of Lesion-Activated Cat OP
In the ECON model of demyelination and remyelination,predivision aOP identifigable by immunostaining for vimentin,are present in fascicles close to early(2~3 days postinjection)lesions(Carroll et al.,1998;Jennings et al.,2002).To compare the abundance and morphology of aOP with normal ON NG2cells,serial 1 μ m sections from peri-lesion ECON blocks were cut and stained alternately with Vim and another glial marker.Vim+aOP counts were then compared with those of NG2cells in 2 μ m paraffign sections of normal cat ON.To ensure that the counting frames(above)enclosed the same area of tissue despite differences in tissue-processing protocols in the two section types,counts were also compared for oligodendrocytes in normal cat ON 1 μ m araldite and 2 μ m paraffign sections.A three-dimension morphological description of aOP was constructed by tracking and imaging individual Vim+cells through serial sections.
RESULTS NG2 Expression Distinguishes a Separate Class of Neuroglia in Cat and Human ON
Frozen sections of cat and human ON reacted with antibody to NG2 revealed a meshwork of fine,tendril-like,cytoplasmic processes throughout the parenchyma,aligned with fibers as well as spanning across them.Blood vessels were strongly delineated due to staining of pericytes(Ozerdem et al.,2002).Neuroglial cells with surface positivity for NG2(NG2cells)were located both alone among the fibers and within rows of other glial cells(see Fig.1).Many ofthefiber-associated NG2cells had elongate nuclei(up to 16 μ m in length)of slightly irregular outline,closely surrounded by the cell membrane and orientated slightly oblique to the parallel as if weaving between adjacent fibers(Fig.1A),an impression strengthened by differentialfocusing through 10 μ m sections.Their long fine processes emanated predominately from the polar regions of the cell body,coursing along nearby nerve fibers before branching and spreading laterally to follow other fibers.In TS ON,processes from morphologically similar NG2cells were observed to encircle fibers,nodes of Ranvier,and other neuroglial cell bodies.Despite extending up to five times the length of the cell body,processes from NG2cells did not appear to overlap.Also among the fibers,less common but prominent NG2cells had regular,ovoid nuclei(~ 7 μ m × 12 μ m)and processes that spanned across the fibers to end amongst glial row cells(Fig.1C).Within these rows,NG2cells had relatively rounded nuclei and processes that arose more equally around their cell bodies,appearing to contact neighboring cells as well as extending out among the fibers(Fig.1D).

Fig.2. NG2cell Olig2 and Olig1 expression in frozen sections.Cat(A,C,D)and human ON(B);(A)combination of NG2 and Olig2 as a mixed primary revealed many Olig2+cell nuclei(arrowheads)including those of NG2cells(arrow)but not of blood vessel-associated NG2+cells(open arrows).B,C:Because of greater Olig2 expression in NG2cells relative to oligodendrocytes,antibody dilution resulted in exclusive staining of NG2cell nuclei(arrows).D:Dual staining for Olig1(brown,arrow)and NG2(gray,arrowheads)demonstrated that nuclear Olig1 expression was a feature of NG2cells.Bars=10 μ m.
Thus,while all NG2cells featured extensive and elaborate processes,there was significant morphological heterogeneity of nuclei and cell bodies.To investigate the possibility of separate populations or subpopulations,antigenic characterization of NG2cells was undertaken after initial trials confirmed their identity separate from the other neuroglial cell types;that is,positivity for NG2 did not overlap with markers foroligodendrocytes(HNK-1,GS,Nogo-A),astrocytes(GFAP,Vim),or microglia(RCA-1,CD68).
All NG2cells Belong to the Oligodendrocyte Lineage
Olig2 is a transcription factor involved in the phenotypic definition of cells in the oligodendroglial lineage(Yokoo et al.,2004).Reaction of frozen sections for Olig2 resulted in nuclear positivity of a proportion of glial cells.To investigate the relationship between NG2cells and the oligodendrocyte lineage,antibodies to NG2 and Olig2 were mixed and reacted as a single NG2Olig2 antibody.All NG2+glial cells were found to have Olig2+nuclei while the blood vessel-associated NG2+cells did not(Fig.2A).Furthermore,the relative intensity of Olig2 staining was such that antibody dilution restricted its detection to NG2cells(Fig.2B,C).Similarly,dual staining for Olig2 and PDGFα R resulted in the restriction of surface staining to cells with intense nuclear Olig2+,supporting identity of the NG2+andPDGFα R+populations.When NG2Olig2 or PDGFα ROlig2 staining was contrasted with glutamine synthetase,allremaining Olig2+cells were confirmed as GS+oligodendrocytes.Reaction for the other oligodendrocyte lineage transcription factor,Olig1,revealed that a minor proportion of the stained cells displayed nuclear rather than cytoplasmic positivity.Dual staining for NG2 demonstrated that the cells with nuclear Olig1 were NG2+(Fig.2D).Together,these reactions generated an antigenic phenotype common to all NG2cells,viz;NG2+/PDGFα R+/Olig2+/Olig+(n)/GS-/HNK-1-/S-100-/GFAP-/Vim-/CD 68-/RCA-1-(Table 3).From this phenotype,NG2cells in frozen sections could now be reliably identified without the need forsurface antigen(NG2 and/or PDGFα R)demonstration on the basis of their staining profile with Olig1 and Olig2,thereby potentiating NG2cell identification in tissue processed for paraffin embedding.

TABLE 3.Antigenic Phenotype of Normal Cat and Human ON Neuroglial Cells
Olig1 and Olig2 Expression Identifies NG2cell Nuclei in Paraffin Sections
Reaction of paraffin sections of cat and human ON for Olig2 and Olig1 resulted in the same expression pattern as that observed in the frozen section material;Olig1 was present as diffuse cytoplasmic staining except in a small number of cells with nuclear positivity;Olig2 was nuclear,with expression strength ranging from weak to the intense positivity characteristic of NG2cells.Comparison of 2-μ m serial sections stained for Olig2 and Olig1,respectively,confirmed that the cells with intense Olig2+co-expressed nuclear Olig1(Fig.3A,B).All remaining Olig2+cells had diffuse cytoplasmic Olig1 and were confirmed as oligodendrocytes by cytoplasmic NoA+or GS+(Fig.3C).
Direct NG2 and PDGFaR Demonstration in Paraffin Sections of Human ON
As final confirmation that the Olig1/Olig2 transcription factorphenotype identified NG2cells,paraffin sections of zincfixed human ON were reacted for NG2 or PDGFα R,either singly or in combination with Olig2.Although proximal process definition was limited compared to frozen sections,fine,tendril-like,NG2+process profiles were present throughout the neuropil against a clean background(Fig.3D).Cell-body NG2+was restricted to the surface of cells with intense nuclear Olig2(Fig.3E).PDGFα R expression was also demonstrable on NG2cells but was less reliable and frequently also stained oligodendrocyte cytoplasm.
Pan-Staining Achieves Differential Identification of All Neuroglia
With NG2cells clearly distinguished by nuclear Olig1 or Olig2,the opportunity existed to simultaneously identify all four classes of neuroglia through multiple antibody-staining protocols(panstaining).For instance,in human ON,antibody to GFAP(astrocytes)as a single mixed primary with Olig2(oligodendrocytes versus NG2cells),contrasted with CD68(microglia),left no neuroglial cells unlabelled(Fig.3F),confirming that Olig2 was expressed by all oligodendrocyte lineage cells;GFAP by all astrocytes;CD68 by all microglia.Such pan-staining also validated simpler regimes that labeled three of the neuroglial classes,assigning the fourth to the coun terstain.For instance,Olig2 contrasted with CD68 stained all neuroglia other than astrocytes;Vim(astrocytes)versus Olig2 left only microglia unstained(Fig.3G)and could be further simplified to a single mixed primary protocol(Fig.3H).Pan-stained sections also revealed detail about the physical relationships between individual neuroglia,highlighting the extensive contact between processes and even cell bodies of the four classes.In particular,some NG2cell nuclei were observed in close apposition to other cell bodies(Fig.3G).Although clearly demonstrable with Olig2-or Olig1-staining,NG2cells could not be identified by nuclearmorphology alone,because theirnuclei,viewed in haematoxylin-stained serial sections,had an unobtrusive appearance and could be similar to those of oligodendrocytes or microglial cells.
NG2cells are Demonstrable in FFPE Human ON
Although sections from human ON with up to 2 weeks of formalin fixation displayed markedly reduced antigen availability compared to the zincfixed specimens,incorporation of AR largely restored the demonstration of intracellular elements of the neuroglialantigenic phenotypes(Table 3).NG2cells were able to be identified by Olig1 nuclear positivity(Fig.3I),by Olig2 alone on the basis of relative intensity(Fig.3J),or by the Olig2+/GS-phenotype(Fig.3K).
Quantification of NG2cells
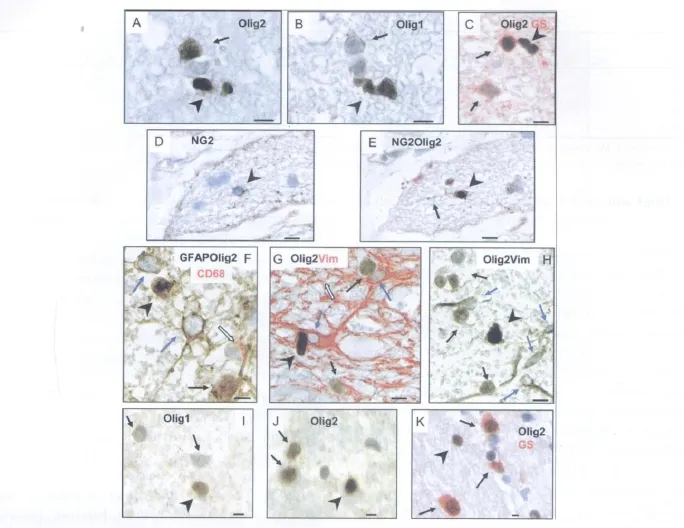
Fig.3. NG2cell identification in paraffin sections.A,B:Serial sections of cat ON stained for Olig2(A)compared to Olig1(B).NG2cells(arrowheads)were identifiable by their intensely Olig2+(A)or Olig1+(B)nuclei,whereas oligodendrocytes(arrows)had less intense nuclear Olig2+(A)and cytoplasmic Olig1(B).C:Dual staining for Olig2(brown)and GS(red)confirmed that all Olig2+cells other than NG2cells were GS+oligodendrocytes.D,E:Serial sections of zincfixed human ON;direct cell surface NG2 demonstration(arrowhead)was achieved either on its own(A)or combined with Olig2(B)where it labeled cells with intensely Olig2+nuclei(NG2+process indicated by arrow).F-H:Pan-staining to simultaneously identify all neuroglia.F:Human ON;GFAP mixed with Olig2 antibody(brown)labeled astrocyte cell bodies and processes(blue arrows)as well as the nuclei of oligoden drocytes(arrow)and NG2cells(arrowhead)while CD68(red)labeled microglial cytoplasm(open arrow),simultaneously identifying all four neuroglial classes.G:Cat ON;Olig2(brown)contrasted with Vim(red)separately identified NG2cells(dark brown,arrowhead),oligodendrocytes(paler brown,arrows),astrocytes(red,blue arrows),and microglia(counterstain only,open arrow).H:Human ON;Olig2 mixed with Vim and reacted as a single antibody distinguished NG2cells(arrowhead)from oligodendrocytes(arrows),astrocytes(blue arrows),and microglia(counterstain,none present).I-K:FFPE human ON;NG2cells(arrowheads)were identifiable by(I)Olig1 nuclear positivity,(J)Olig2 alone,separated from oligodendrocytes(arrows)on the basis of relative intensity,or(K)with dual staining for GS to separately identify oligodendrocytes(Olig2+GS+;arrows).Bars=5 μ m.
NG2cells were counted as cells per unit area(mm2)and in comparison with oligodendrocytes,astrocytes,Olig2+oligodendrocyte lineage cells,microglia,total macroglia,and total neuroglia,in both standard fields(0.2 mm2)and totalled across full nerve faces(see Methods section).
Cat ON(Table 4)
In cat ON,there were 22 NG2cells per mm2compared to oligodendrocytes(272)and astrocytes(210).NG2cells comprised 8%of the oligodendrocyte lineage(total Olig2+cells),4~5%of the macroglia(oligodendrocytes,55%;astrocytes,41%),and 3%of the total neuroglial complement(oligodendrocytes,47%;astrocytes,37%;microglia,13%).The ratio of NG2cells to oligodendro cytes was 1∶12.The same ratio resulted for frozen section counts of NG2+neuroglia in comparison with GS+oligodendrocytes.
Cat SC(Table 5)
To investigate whether the ratio of NG2cells to oligodendrocytes was restricted to ON or was more generally representative of CNS tracts,the samecounting protocol was applied to identically processed cat SC sections(Table 5).Counts in SC WM approximated those in ON(274 compared to 295 oligodendrocyte lineage cells per mm2).Although absolute counts in gray matter(GM)exceeded those in WM(34 compared to 23 NG2cells per mm2and 434 compared to 251 oligodendrocytes per mm2),the ratio between NG2cells and oligodendrocytes was similar in GM,WM,and ON at 1∶13,1∶11,and 1∶12,respectively.
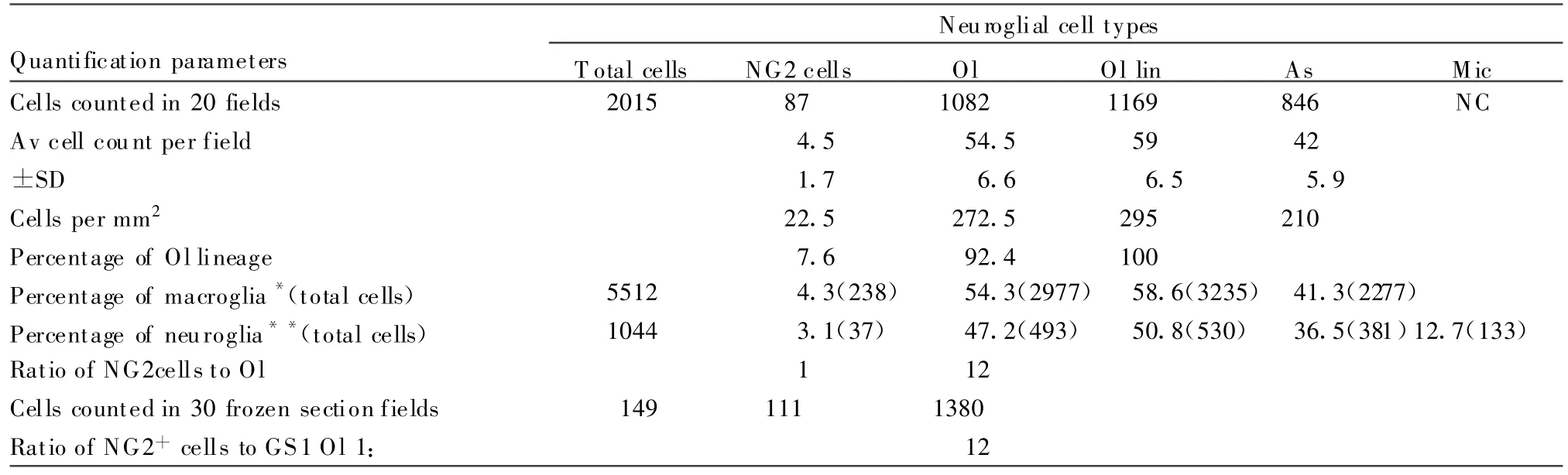
TABLE 4.Quanti ficationof Neuroglial Cell Types in Paraf fin and Frozen Cross-Sections of Cat ON

TABLE 5. Quantif icationof Oligodendrocyte Lineage Cells in Cat ON,White Matter,and Gray Matter Regions of Cervical Spinal Cord
Human ON(Table 6)
In human ON,there were 35 NG2cells per mm2compared to oligodendrocytes(340),astrocytes(205),and microglia(90).NG2cells comprised 5%of the total neuroglia(oligodendrocytes,51%;astrocytes,31%;microglia 13%),6%of the macroglia(oligodendrocytes,58%;astrocytes,36%),and 10%of the oligodendrocyte lineage.T he ratio of NG2cells to oligodendrocytes was 1∶10.
E f f ect of gender on human oligodendrocyte lineage abundance(Table 6).In rodent CNS WM,a significant gender difference in oligodendrocyte abundance has been reported,with oligodendrocyte counts~30%higher in male mice and rats than in females(Cerghet et al.,2006,2009).To investigate whether the same might apply in human WM,we compared oligodendrocyte lineage abundance between female and male human ONs.In standard field counts of Olig2-stained sections from similar age males and females,the average number of oligodendrocyte lineage cells was almost identical(female,76 cells;male,75 cells).Counts were not carried out in cat material,because males have been neutered.
Ef fect of age on human oligodendrocyte abundance(Table 6).In rhesus monkey ON,inherent age-related demyelination has been reported to result in an up to 40%increase in oligodendrocytes,generated to remyelinate the shorter internodes(Sandell and Peters,2002).To investigate the effect of ageing on oligodendrocyte abundance in human ON,we compared counts(averaged from five standard fields)in the youngest human,an 18-year-old female,with that in the oldest,an 82-year-old male(Table 1).Notwithstanding the lackof a demonstrable gender difference(above),counts were also made in relatively aged female ON(59 years).Although there were slightly more oligodendrocytes on average in the standard count area of the older female(72 cells)and the oldest individual(71 cells)compared to the youngest(68 cells),all values overlapped within the range of one standard deviation.However,these counts did not take into account factors such as the thicker interfascicular septa in the aged ONs that effectively decreased the countable portion of fields.
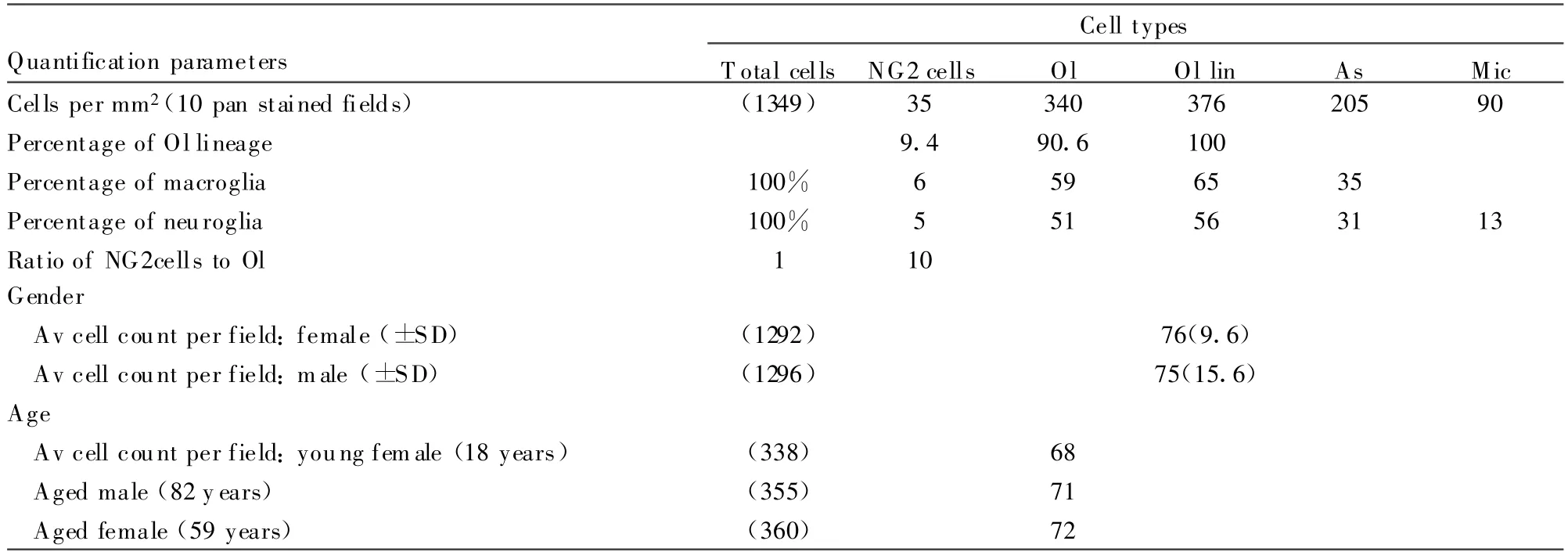
TABLE 6.Quantification of Neuroglial Cell Types in Paraf fin Cross-Sections of Human ON
NG2cells Activate in Response to Local Demyelination
With a ratio of one NG2cell for every 10~13 oligodendrocytes,it was important to investigate whether all NG2cells,or only a proportion,could respond as OP in the event of local oligodendrocyte loss.As set out in the Methods section and Table 7,we assessed this by comparing counts of predivision activated(Vim+)OP in 1 μ m araldite sections of cat ON containing an early demyelinative lesion(Fig.4C)with NG2cells in 2-μ m paraffin sections of normal cat ON(Fig.4B).The resultant similarity(22 aOP/mm2vs.22.5 NG2cells/mm2)suggested total or near total NG2cell activation as OP and was consistent with NG2cells comprising a single neuroglial class.
The ability to stain aOP cytoplasm with antibody to Vim also permitted their morphological description.Individual predivision,and therefore in situ aOP,tracked and imaged through serial 1 μ m sections,had a uniform morphology featuring a large(~ 8 μ m diameter)ovoid nucleus enclosed within a regular cell body from which emanated one principal and one minor polar process(Fig.4D).This relatively simple morphology was in marked contrast to the complex,process-rich morphology of unactivated NG2cells visualized in normal ON frozen sections(Fig.4A).
DISCUSSION
Although surface staining for NG2 is considered the hallmark for identifying OP in the adult CNS,controversy exists as to whether NG2+neuroglia comprise functional subtypes,particularly with respect to capacity for oligodendrocyte generation(Wigley and Butt,2009).The importance of resolving this issue is highlighted by a current focus in MS research on enhancement of endogenous repair,which is premised on the human CNS possessing a robust remyelination capability sponsored by abundant OP(Dawson et al.,2003).In this study,NG2cells have been identified,morphologically characterized,and quantified in adult cat and human ON through correlation of fixation sensitive surface markers with newly described and established intracellular neuroglial markers.In paraffin sections,NG2cells were uniquely identifiable by intense staining for Olig2 and/or nuclear Olig1.Although they were shown to belong to a single class of neuroglia,all with OP capability,quantification revealed that NG2cells were not abundant,comprising less than 5%of total neuroglia and,importantly,less than 10%of oligodendrocyte lineage cells.
Initial identification was in frozen sections,where cell membrane NG2 expression outlined cell bodies and fine processes that extended throughout the neuropil,intimately associating with the outer surfaces and nodal features of fibers as well as with other neuroglia.Despite their collective distinction from astrocytes,microglia,and oligodendrocytes,NG2cells displayed significant morphological heterogeneity(see Fig.1).Did this refiect the functional heterogeneity proposed for NG2cells,with the more common,elongate,fiber-associated cells equating to “synantocytes” and only the rarer,ovoidcells comprising OP,or were the morphological differences a response to subtle environmental variation as proposed for“polydendrocytes”Because synantocytes are considered to be differentiated,postmitotic cells,our findings that all NG2cells expressed PDGFα R,Olig1,and Olig2 are in line with the concept of a single“polydendrocyte” cell type within the oligodendrocyte lineage.Proliferative progenitor capability was supported by nuclear expression of Olig1(Arnett et al.,2004;Kitada and Rowitch,2006;Xin et al.,2005)and by the observation in the ECON model that aOP abundance matched that of normal ON NG2cells.Importantly,although all appear to possess OP capability,our observations of NG2cells'intimate relationships with nerve fibers and the other neuroglia support the contention that they are not simply immature or“resting”oligodendrocyte precursors.Given their morphological heterogeneity and complexity,it is possible that functional NG2cell subtypes exist,potentially separable in future studies on the basis of differential expression of additional oligodendrocyte lineage transcription factors(Fu et al.,2009;Kitada and Rowitch,2006).
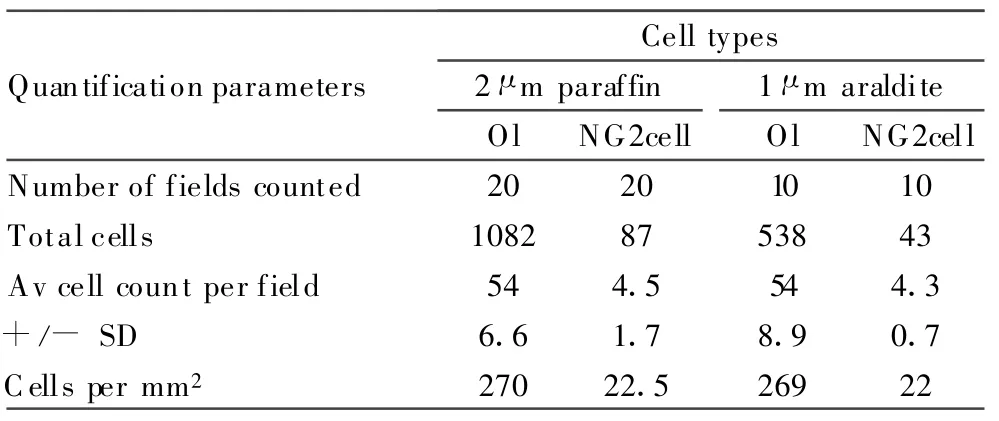
TABLE 7. Quantification of Activated OP in the Fascicles Surroundingan Early Demyelinating Lesion Compared with NG2cellsand Oligodendrocytes in Normal Cat ON
The relative simplicity of the ON facilitated the identification,characterization,and quantification of NG2cells through the ability to differentially immunostain all neuroglia,both in serial cross-sections and within single pan-stained sections.Correlation of frozen section surface staining(NG2 and PDGFα R)with nuclear transcription factor expression led to the identification of NG2cells in paraffin sections on the basis of nuclear Olig1 and/or intense Olig2,as part of the overall intracellular antigenic phenotype;Olig2++/Olig1(nuclear)+/GS-/NoA-/HNK-1-/GFAP-/Vim-/CD68-/RC A-1-(Table 3).T he subsequent application of this phenotype to FFPE human ON(Fig.3I-K)provides the prospect of reliable NG2cell identification in conventionally prepared human specimens.Importantly,even in perfused or short pmi tissue,antigenic phenotyping was essential for identification,because NG2cells could not be identified on the basis of morphology alone,in line with previous observations(Peters et al.,2008).Additionally,as pmi increased,NG2cell nuclei tended to appear rounded and uniform,which could lead to the erroneous impression that NG2cells were of similar morphology to oligodendrocytes(Wilson et al.,2006).
Accuracy of quantification was attained by three approaches.First,identification of each neuroglial cell type through its unique antigenic phenotype ensured that no cells were left unidentified within quantification fields(Table 6).To date,as suggested by a recent review including a table collating NG2+cell counts,this study may be the only one to have quantified and compared all component neuroglial classes of a CNS tract(Nishiyama,2007).Second,counting was of a single layer of cells(2 ~ 3 μ m sections)rather than multiple layers requiring correction factors for interstudy comparison(Wolswijk,2002).Third,results were expressed as relative percentages in addition to cells per unit area.As has been pointed out,“more important than the absolute size of the NG2-expressing cell population,is its relativity to other glial cell types and in particular,in terms of the success and relative speed of repair,the ratio of progenitors to oligodendrocytes”(Dawson et al.,2003).
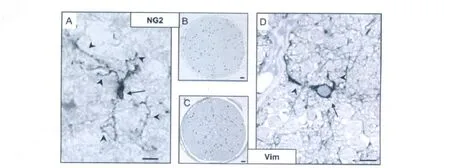
Fig.4. Comparative morphology and abundance of NG2cells and aOP.A,B:Normal cat ON;(C,D)peri lesion cat ON(2 days postinjection).A:An NG2cell in normal ON features an elongate,irregular cell body(arrow)and long,fine,branching processes(arrowheads).D:In marked contrast,following NG2cell activation in response to nearby demyelination,an aOP features a regular,ovoid cell body(arrow)and relatively short,simple,polar processes(arrowheads).B,C:Whole face sections overlaid with dots representing counts of individual NG2cells(B)and aOP(C)illustrate the similarity between pre and postactivation abundance,supportive of majority NG2cell activation.Bars=10μ m(A,D);0.1 mm(B,C).[Color figure can be viewed in the online issue,which is available at www.interscience.wiley.com.]
Human ON NG2cells were quantified at 35 per mm2in comparison with astrocytes(205),microglia(90),and oligodendrocytes(340).NG2 cells comprised 5%of the total neuroglia,6%of the macroglia,and 10%of the oligodendrocyte lineage,a ratio to oligodendrocytes of 1∶10.In fact,the OP:oligodendrocyte ratio was remarkably consistent between human and cat ON(1∶12)as well as between cat WM(1∶11)and GM(1∶13).An implication of this numerical imbalance between OP and oligodendrocytes is that to fully repair demyelination,OP recruitment would need to occur from a larger area of the surrounding unaffected WM than the lesion itself and/or necessitate several rounds of division.However,in experimental demyelination,local OP recruitment is limited to the immediate lesion surround,with further restrictions due to ageing(Franklin et al.,1997;Rist and Franklin,2008;Shen et al.,2008;Sim et al.,2002),while OP division has been reported to be symmetric,whereby both daughter cells commit to remyelination(Carroll et al.,1998;Keirstead and Blakemore,1999;Keirstead et al.,1998).Therefore,even with all NG2cells having OP capability,our findings predict the potential for repair insufficiency due to OP depletion in the event of large-scale,protracted or repeated demyelination,at least in tracts lacking the potential for replenishment from germinal zones(Aguirre et al.,2007;Small et al.,1987).That this is the end point for many ON MS lesions is suggested by a study of the ONs from 18 MS autopsy cases where gliotic,chronically demyelinated lesions were described in 35 of 36(Ulrich and Groebke-Lorenz,1983).
By achieving reliable identification and quantification of NG2cells,this study has demonstrated that all NG2cells in the ON are OP despite their morphological complexity.However,the finding that OP are not abundant,particularly relative to oligodendrocytes,may have implications for efforts to enhance endogenous repair in MS.
Dr Jennings was in receipt of a PhD scholarship from MS Research Australia.The authors thank Dr Ghosh for cat optic nerve samples;the Lions Eye Bank for human optic nerve samples;Dr Stallcup for antibodies to NG2 and PDGFα R;Professor Stiles and Dr Alberta for antibodies to Olig1 and Olig2 and Dr Everett for HNK-1 antibody.
